
U okviru teksta “Uvod u evoluciju”, objašnjen je proces nastanka genetskih informacija kroz duplikaciju praćenu mutacijama. No, ovo nije jedini način na koga informacije mogu nastati, i vredi pomenuti par dodatnih procesa.
Viralni transfer je proces u okviru koga retrovirusi unose nove gene u genom organizma koga su napali.
Većina virusa deluje tako što u ćeliju ubaci svoj genetski materijal, koji onda preuzme kontrolu nad ćelijom. Umesto svojih normalnih procesa, ćelija počne da proizvodi nove kopije virusa. Na kraju, ćelija umire i raspadne se, oslobađajući nove viruse, koji sada mogu da napadnu druge ćelije.
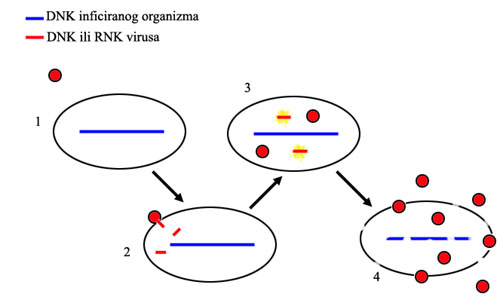
Tok virusne infekcije: 1. virus se približi ćeliji, 2. virus se spoji sa ćelijom i u nju ubaci svoj genetski materijal, 3. geni virusa postanu aktivni i ćelija na osnovu njih proizvede još virusa, 4. ćelija umire i raspada se, oslobađajući gomilu novih virusnih čestica.
Retrovirusi idu korak dalje: oni ne samo što ubace svoj genetski materijal u ćeliju, već ga zatim i integrišu u genom “domaćina.” Rezultat je da ćelija ne može da razlikuje virusne gene od svojih sopstvenih. Virus ne mora čak ni da bude aktivan, može da se ućuti i da prosto čeka, što mu omogućava da prebrodi imunu reakciju unutar tela. Onda, u nekom budućem trenutku, geni se aktiviraju, i preuzmu kontrolu nad ćelijom; ćelija zatim umre proizvodeći gomilu novih virusa. Najpoznatiji retrovirus danas je HIV, virus koji uzrokuje sidu.
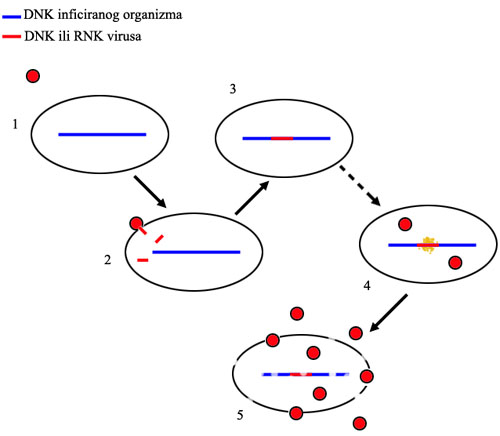
Tok retrovirusne infekcije: 1. retrovirus se približi ćeliji, 2. retrovirus se spoji sa ćelijom i u nju ubaci svoj genetski materijal, 3. geni virusa se utope (integrišu) u genom organizma, i tu mogu da ostanu mesecima ili godinama bez aktivnosti, 4. geni postanu aktivni i ćelija na osnovu njih proizvede još virusa, 5. ćelija umire i raspada se, oslobađajući gomilu novih virusnih čestica.
Kako ovi virusi utiču na evoluciju? Pa, povremeno će retrovirus inficirati neku od semenih ćelija (spermatozoid ili jajnu ćeliju). Time će potomci inficirane jedinke naslediti virusne gene zajedno sa svim ostalim.
Od ove tačke, mogu se desiti tri stvari. U prvom slučaju, ako su virusni geni netaknuti, virus će se aktivirati i potomci koji su nasledili gene će takođe naslediti i virusnu bolest. Ona će ih oslabiti ili ubiti, i time eliminisati ovu genetsku kombinaciju iz populacije. U drugom slučaju, virusni geni mogu biti oštećeni mutacijama (veliki broj mutacija biva unesen u toku mejoze koja proizvodi semene ćelije). U ovom slučaju, ovi geni postaju deo “DNK đubreta”, i postaju neupotrebljivi. U trećem slučaju, virusni geni bivaju malkice oštećeni, ali su i dalje aktivni. Šta se u ovom slučaju događa, zavisi od mesta u genomu u koje se virus ubacio.
Ako se virus ubacio u genom na mesto na kome virusni geni štete organizmu, ili na mesto na kome nemaju uticaja, oni će biti eliminisani iz populacije, ili će prosto propasti (kao što je objašnjeno u tekstu o selektivnim pritiscima), ili će se pretvoriti u transpozone (kao što je objašnjeno u tekstu o nekodirajućim tragovima u DNK). No, ako se virus ubacio na mesto na kome geni mogu da vrše neku pozitivnu funkciju, oni postaju aktivan deo genoma organizma. Ovo je veoma retka stvar, ali povremeno može imati veoma značajne posledice.
Primer: ERV-3. Prvi sisari su bili monotreme: bića koja su imala mlečne žlezde, ali su pritom legla jaja kao i njihovi preci (dva predstavnika ove grupe postoje i danas: kljunar i ehidna). Jaja monotrema imaju ljusku i jajnu membranu. U daljoj evoluciji, ljuska jajeta je omekšala, stopila se sa jajnom membranom, i postala ono što danas zovemo placenta: membrana koja okružuje novonastali embrion.
U jednom trenutku u toku ove evolucije, predak svih placentalnih sisara je postao zaražen retrovirusom. Između ostalih gena, ovaj virus je nosio gen koji potiskuje imuni sistem, što je omogućavalo virusu da se lakše odupre imunim napadima. Kod jedne jedinke, virus se integrisao u semene ćelije, i to u delu DNK koji je aktivan u placentalnom tkivu.
No, u ovom slučaju, rezultat integracije je bio da se ovaj gen aktivira u placentalnom tkivu: tkivu koje okružuje embrion i štiti ga od imunog sistema majke. Prisustvo ovog gena je imalo pozitivan efekt na reprodukciju, smanjenje šanse da dođe do imunološkog odbacivanja fetusa. Samim tim, ovaj gen je potpao pod prirodnu selekciju, i postao je deo opšteg placentalnog genetskog sistema. Danas svi placentalni sisari i dalje nose ostatak ovog virusa u svom genomu; svi drugi delovi su degenerisali, ali ovaj imunosupresivni gen je ostao aktivan, prisutan, i pod konstantnim selektivnim pritiskom
Mešanje eksona je drugi važan mehanizam kojim mogu nastati novi geni. Radi razumevanja, međutim, potrebno je poznavanje jednog važnog aspekta genetike eukariota (grupe živih organizama u koje spadaju sve biljke i životinje).
Naime, bakterije i arheje uglavnom poseduju “jednodelne” gene: svaki gen je “zapisan” od početka do kraja u genomu organizma. Eukariote, sa druge strane, imaju gene koji su podeljeni u aktivne delove (“eksone”) i neaktivne delove (“introne”). U toku aktivacije gena, ceo gen biva iskopiran u RNK, nakon čega introni bivaju isečeni, a eksoni spojeni u celinu. Ovo omogućava da isti gen proizvede više različitih vrsta proteina (koji se sastoje od različitih kombinacija eksona) – u nekim slučajevima, više stotina, hiljada ili čak desetina hiljada verzija (jedan šampion na ovom polju je gen DsCam, iz vinskih mušica: iz jednog gena, biva proizvedeno čak 38,000 različitih proteina. Ovo strahovito povećava raznovrsnost genoma: ljudski genom se sastoji od oko 20-25 hiljada gena, ali usled varijacije u eksonima proizvodi oko 250,000 različitih proteina.
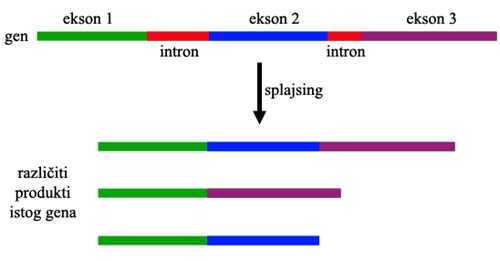
Pojedini funkcionalni delovi gena su često sadržani u pojedinačnim eksonima (ili grupama eksona). Više različitih genetskih procesa (na primer, hromozomalno ukrštanje) mogu dovesti do situacije u kojoj deo jednog gena završi u okviru nekog drugog. Rezultat ovoga je protein koji se sastoji od svih prethodnih delova, ali sada ima još jednu ili više dodatnih funkcionalnih celina. Isto tako, eksoni različitih gena mogu postati duplikovani i kombinovani u potpuno nove gene.

F – f-modul, FN2 – fibronektin modul tip II, G – modul faktora rasta,
K – kringl modul, SP – proteolitički domen
Kao jedan konkretan primer (od hiljada postojećih), na gornjoj slici su prikazana četiri proteina koja su nastala mešanjem eksona. Kopiranjem i mešanjem različitih domena, stvorene su četiri različite proteaze (enzimi koji seku druge proteine pod određenim uslovima), koje imaju četiri različita uslova aktivnosti i četiri različite mete.