
- Uvod – šta su to prelazni fosili
- Razvoj višećelijskih organizama i Kambrijska eksplozija
- Razvoj kičmenjaka
- Od riba do vodozemaca
- Od vodozemaca do reptila
- Od reptila do sisara
- Od reptila do ptica
- Kratak pregled evolucije biljaka
- Žirafe, kitovi, konji i druge napomene
Čitaocima se preporučuje da, ako već nisu, prvo pročitaju tekst “Uvod u Evoluciju”, pre nego što počnu sa čitanjem ove analize..
Kako fosili nastaju u prirodi?
Svi znamo šta se događa sa mrtvim životinjama: njihovo telo se raspada, trunući dok od njega ne ostane praktično nikakvog traga. U veoma retkim uslovima – otprilike jednom u svakih nekoliko miliona slučajeva smrti – telo životinje se može naći u uslovima koji omogućavaju procese fosilizacije. Ovi procesi najčešće neće očuvati ništa od početne organske materije tela; telo u potpunosti istruli, ali iza sebe ostavi trag, otisak svoje strukture u okamenjenim mineralima. Telo nestaje, a nama ostaje oblik tog tela očuvan u kamenu. (U još daleko ređim slučajevima, doslovno jednom u više milijardi, mali tragovi organske materije mogu ostati delimično očuvani kroz veoma duge periode vremena, uglavnom u veoma izolovanim delovima tela kao što je unutrašnjost debelih i snažnih kostiju.)
Šta je cilj ovog teksta?
Dok se tekst “Evolucija Čoveka” bavi fosilima koji pokazuju razvoj ljudi od primata, ovaj tekst će se koncentrisati na opisivanje evolutivnog puta od primitivnih jednoćelijskih organizama do primata. Ovo znači da će najveći deo evolutivnog stabla – evolucija riba nakon što su se od njih odvojili preci vodozemaca, evolucija vodozemaca nakon što su se od njih odvojili preci reptila, itd. – biti samo usputno pomenut.
Razlog za ovo je veličina posla: u toku zadnjih par vekova, paleontolozi su iskopali bukvalno milione fosila, uključujući tu i desetine hiljada raznih prelaznih vrsta. Mogli bi smo ovde napisati čitave knjige, pa da opet ne uspemo da obradimo ni pola poznatih fosila.
Čitaoce koji žele da nauče više o ovim drugim granama evolutivnog razvoja upućujemo da pogledaju u sekciju sa linkovima, gde se nalaze liste linkova na mnoge stranice koje sadrže daleko veću količinu detalja.
Procesi fosilizacije kroz koje se živi organizmi mogu ovako “okameniti” su dobro poznati modernoj nauci, i možemo ih posmatrati uživo u prirodi. Pažljivim kopanjem se mogu u tlu naći ostaci koji su u svim stepenima fosilizacije, od potpuno svežih, preko polu-fosilizovanih, do fosila kod kojih je tkivo potpuno zamenjeno mineralima
Najčešći oblik fosilizacije su otisci. Organizam potone u blato ili mulj, koje se zatim osuši. Organizam istrune potpuno, a u sledećem naletu kiše novi sloj peska ili blata prodre u rupu koja je tako nastala. U toku mineralizacije sedimenta kojom blato postaje kamen, ova dva sloja blata ostaju odvojeni. Kasnije nastali kamen se može po ovom procepu odvojiti, ostavljajući nam otisak davno istrulelog tela. Ovo povremeno proizvodi i takozvane endokaste: organizam koji je već počeo da trune potone na dno, i u njegove unutrašnje šupljine prodre blato. Kada se to blato učvrsti, ostaje nam otisak unutrašnje šupljine organizma.
Slede tri vrste mineralizacije. Prva vrsta je permineralizacija, u kojoj organizam potone u sedimente veoma brzo, nakon čega mineralima bogata voda iz okoline prodre u ostatke i precipituje mikrokristale; organski delovi organizma se u ovim uslovima polako raspadnu, ali mineralizovana struktura ostaje okamenjena u obliku istrulelog tela.
Druga vrsta mineralizacije je autigena mineralizacija. Ovo se događa samo u veoma specifičnim uslovima, mada oni nisu posebno retki. Organizam živi u jednom sloju vode, i nakon smrti potone u dublje slojeve, koji su mineralno različiti. Telo organizma pada u te donje slojeve, i tamo postaje nukleus za naglu mineralizaciju oko sebe: doslovno se oko organizma u roku od nekoliko sati ili dana formira mineralni oklop. Organizam u ovakvoj noduli na kraju potpuno istruli i pretvori se u prah, ali oko njega ostane jasan otisak oblika tela. Ovaj oblik mineralizacije se može danas uživo posmatrati u mnogim podvodnim pećinama.
Poslednji oblik mineralizacije, adpresija, nastaje zatrpavanjem organizma pod pritiskom. Recimo, odron kamenja odnese deo rečne obale, prilikom čega biljke padnu na dno i onda budu ugurane u mulj; kamenje stoji odozgo i pritiska celu stvar. U ovom slučaju može doći do kompleksne serije reakcija kojim molekuli samog živog bića reaguju sa okolnim mineralima, ostavljajući spljošten otisak unutar sedimentnog sloja. Ova vrsta mineralizacije se obično dešava kod biljaka, čiji sastavni materijal lignin predstavlja posebno dobru osnovu za potrebne hemijske reakcije.
Konačno, tu su fosili koji nastaju bioimuracijom. Ona se dešava kada egzoskeletalni organizam prekrije ostatke životinje. Recimo, koralne larve mogu da prekriju kosti nedavno uginule ribe; u periodu truljenja kostiju, korali izrastu dovoljno da se povežu jedni sa drugima, i u koralu ostaje šupljina u obliku otiska istrulelih kostiju.
U kakvom stanju se nalazi većina fosila?
Najveći deo fosila koje imamo su…mrvice. U stenama se pod mikroskopom jasno mogu videti fosilizovani parčići žvakanih kostiju. Postoje čitavi geološki slojevi, metrima duboki, koji nisu ništa drugo do okamenjene naslage kokolita, ribljeg izmeta u kome se nalazi gomila sažvakanih ostataka drugih organizama. Ovakve naslage se u svetu mere doslovno stotinama miliona tona, i predstavljaju otpatke koji su se gomilali kroz desetine ili stotine miliona godina.
Čak i čuvene fosilne naslage, kao što su one iz Kambrijumskog doba, sadrže uglavnom ovakve komade i mrvice. Praktično nijedan of fosila trilobita nije ostatak naglo zatrpane životinje, već se na njemu mogu naći tragovi napada drugih bića (mnogim trilobitima je donji deo tela potpuno odlomljen, što je napadač uradio da bi iz ljušture mogao da isisa trilobitovo meso). Od preostalih fosila, velika većina je delimična: pojedinačni odlomljeni udovi, pojedine kosti (često sa tragovima zuba na sebi).
Tek veoma mali deo fosila čine skeleti celih životinja. Oblasti u kojima se ovakvi fosili najčešće nalaze, kao što su prastara priobalna područja ili močvare, predstavljaju veliko naučno blago. Iz ovog razloga, mi čak i danas imamo kompletne fosile samo malog broja svih vrsta koje su nekada postojale.
Najređi (i najbolji) od svih fosila su oni koji nastaju naglim zatrpavanjem. U svakih milion fosila retko možemo naći čak i jedan jedini koji je nastao na ovakav način; ali kada nađemo takve fosile, oni su spektakularni. Recimo, pogledajte fosil pernatog dinosaurusa Mei long, koji je ostao zatrpan kada se njegova jazbina urušila dok je on u njoj spavao:

Vulkanske erupcije takođe često proizvode velike i upečatljive naslage fosila. U zalivu Fundi, recimo, postoje formacije koje su debele skoro tri kilometra, a koje se sastoje od slojeva okamenjenih šuma. Naime, u blizini je bio aktivan veoma snažan vulkan, koji bi šume zatrpao u pepelu. Stabla bi se tako okamenila, a na površini iznad njih bi polako izrasla nova šuma. Sledeća erupcija bi zakopala novu šumu, i tako nekoliko stotina puta. U okviru šume su okamenjeni i fosili mnogih drugih živih bića iz tadašnjeg ekosistema.
Kakve fosilne nalaze predviđa teorija evolucije?
Procesi evolucije su detaljno objašnjeni u tekstu “Uvod u Evoluciju”; ali u kontekstu prelaznih fosila, vredi ponoviti jednu važnu činjenicu. Evolucija se ne odvija kroz nagle mutacije, niti kroz velike promene u strukturi organizma. Naprotiv, ona je gradualan, postepen proces u kome se postepeno menja čitava populacija vrste.
Katastrofalne mutacije se dešavaju u svakoj generaciji: određen broj novorođenih jedinki bilo koje vrste će imati neku veliku i tešku promenu u svom genetskom kodu. Najveći deo ovih mutacija će proizvesti probleme sa ćelijskom strukturom ili biohemijom, dovodeći do bolesti ili smrti. Ali jedan deo ovakvih mutacija će proizvesti velike deformacije u strukturi tela. Takve mutacije u okviru prirode uvek bivaju ili smrtonosne, ili sprečavaju da organizam normalno živi i da ostavi normalno potomstvo.
Otud, životinje koje dožive ovakve mutacije nikada neće učestvovati u procesu evolucije. One će biti eliminisane iz populacije, i njihovi geni neće biti predati sledećoj generaciji. Takođe, ovakve životinje će jako retko ostaviti bilo kakve fosile. Fosilizacija je efektivno slučajan proces. Ako se od sto miliona pripadnika neke vrste ribe, na primer, fosilizuje ukupno pet ili šest primeraka, koje su šanse da će tih pet ili šest “ubosti” baš neku od malobrojnih deformisanih riba?
Samim tim, prelazni fosili o kojima govorimo u ovom tekstu nisu primerci deformisanih organizama, niti predstavljaju jedinke koje su doživele neku naglu, katastrofalnu promenu. Svaki fosil u ovom tekstu predstavlja okamenjeni primerak organizma koji je bilo deo čitave vrste identičnih (ili veoma sličnih) organizama. Ostaci ovih vrsta nam, kroz svoj vremenski redosled i svoju ugnježdenu hijerarhiju, pokazuju kako nagomilavanje malih promena iz generacije u generaciju dovodi do prelaska iz jedne vrste u drugu.
Tekst koji sledi objašnjava fosile od prvih jednoćelijskih organizama, sve do direktnih predaka ljudi. Ali ako ste nestrpljivi, i želite da vidite najbolje primere postepene promene vrsta, predlažemo vam da odmah pogledate prelaz od riba do vodozemaca, ili prelaz od dinosaurusa do ptica.
Koji redosled fosila možemo da očekujemo?
Veoma je važno razumeti da fosilni zapis ne predstavlja linearan niz, od direktnih predaka do direktnih potomaka. Iz morfološke analize se može jasno videti koje vrste su srodne, i može se videti njihovo mesto u ugnježdenoj hijerarhiji. Ali ako imamo fosil starije vrste, i srodni fosil mlađe – mi ne možemo znati da li je starija vrsta direktan predak (“otac” ili “deda”) kasnije vrste, ili je u pitanju “rođak” druge vrste (“stric” ili “ujak”). Bez mogućnosti DNK analize, samo na osnovu podataka iz oblika kostiju, takve detalje je prosto nemoguće uspostaviti.
Otud, kada neki biolog kaže da je “Arheopteriks predak ptica,” on time ne tvrdi da je Arheopteriks doslovno izlegao jaje iz koga su se rodili preci današnjih ptica. Fosil Arheopteriksa je ostatak vrste koja je bila blizak rođak vrstama iz kojih jesu nastale ptice. Arheopteriks je predak ptica u istom smislu u kome je brat vašeg pradede vaš predak – bio je bliski rođak, deo porodice iz koje potiču vaši geni.
Razvoj vrsta takođe nije linearan proces. Vrste se neprestano dele na grupe i podgrupe, od kojih svaka evoluira zasebno u svom nezavisnom pravcu. Samim tim, predačka vrsta ne mora da izumre da bi iz nje nastala neka nova. Da bi smo ovo pravilno objasnili, korisno je početi od jednog prostog teorijskog primera.
U davnoj prošlosti, ljudi su pripitomili vukove, i kroz veštačku selekciju počeli da upravljaju njihovom daljom evolucijom. Između ostalog, razvijene su rase velikih pasa, mnogo većih od vukova, među kojima su bili i mastifi. Iz mastifa su zatim razvijeni moloseri, a iz molosera bernandinci.
Pritom, nijedna od ovih rasa nije izumrla. I vukovi, i mastifi, i moloseri i bernandinci postoje i dan danas. Razvoj ovih pasa nije bio linearan, već razgranat:
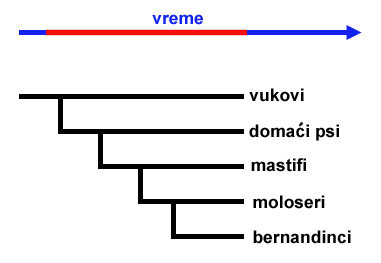
Zamislimo, međutim, da se desi nekakva katastrofa, koja istrebi sve pse na svetu osim bernandinaca. I u dalekoj budućnosti, mnogo milenijuma nakon ove katastrofe, paleontolozi pokušavaju da rekonstruišu kako su bernandinci nastali. Kopajući kroz tlo, oni nađu nekoliko fosilnih ostataka (označeni plavim tačkama):
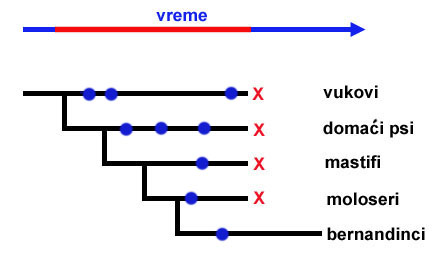
Ovi fosilni ostaci predstavljaju veoma jasnu sekvencu koja pokazuje kako se kostur vuka polako, korak po korak, promenio i postao kostur bernandinca. Možemo videti kako se prvo razvilo veliko telo, pa zatim se promenio oblik lobanje, i konačno vidimo niz promena koje su dovele do samih bernandinaca. Ali pritom, imamo jednu jasnu komplikaciju: nađeni fosili su pogrešnih starosti.
Pogledajte gornju sliku: pošto su vukovi postojali dugo nakon nastanka bernandinaca, nađeni su fosili vukova koji su i mlađi i stariji od fosila prvih bernandinaca. Fosil mastifa je stariji, ali je fosil molosera stariji od mastifovog! Iako je sekvenca jasna sa stanovišta oblika kostiju, datumi se neće slagati.
Da li to znači da ti budući paleontolozi nikada neće moći da rekonstruišu nastanak bernandinaca? Ne. Pošto mi ne možemo da kontrolišemo koja će biti starost fosila koje nađemo, mi uvek moramo očekivati da će u okviru relativno kratkih vremenskih perioda njihova starost biti “zamućena.” Zašto kažemo “relativno kratkih?” Pa, pogledajmo kako situacija izgleda kada nastanak bernandinaca pogledamo iz više perspektive:
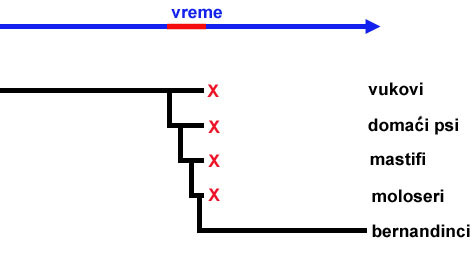
Dugo vremena su postojali vukovi, i nije bilo bernandinaca. Onda imamo relativno kratak period u kome postoje razne međuvrste, i zatim imamo dug period u kome postoje bernandinci. Iako verovatno nikada nećemo naći sve fosile poređane po pravom redu, možemo sa visokim nivoom sigurnosti da zaključimo da su bernandinci nastali od vukova.
Uzmimo sada jedan realan primer: kako su od dinosaurusa nastale ptice.
U sedmom poglavlju ovog teksta je objašnjen jedan mali deo ogromnog broja fosila koji nam pokazuju kako su korak po korak nastali pernati dinosaurusi, kako su od njih nastali pticoliki dinosaurusi, i konačno kako su nastale ptice.
Ali u kontekstu pitanja kojim se ovde bavimo, pogledajmo samo jednu uprošćenu shemu evolucije ptica:
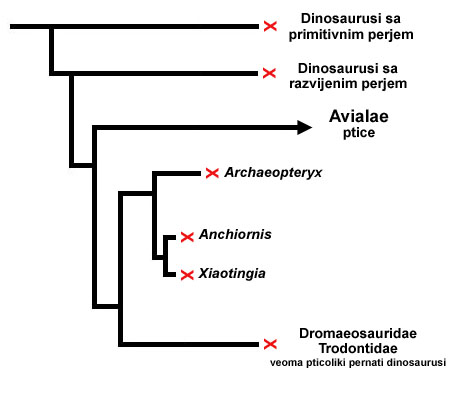
Vrsta dinosaurusa koja je razvila primitivne perolike strukture se podelila na mnogo vrsta, među kojima je nekoliko razvilo kompleksnije perje. Ali vrste sa primitivnim perjem su opstale i postojale su sve do istrebljenja dinosaurusa pre 65 miliona godina. Od vrsta sa kompleksnim perjem su nastale mnoge druge, tako da nalazimo fosile pernatih dinosaurusa (koji nisu ptice) sa najrazličitijim oblicima i strukturama tela. Među ovim vrstama su bili i preci pticolikih dinosaurusa. Od ove grupe su se razvile sa jedne strane veoma pticoliki dinosaurusi (uključujući tu i čuvenog Arheopteriksa), a sa druge preci modernih ptica. Arheopteriks, otud, nije bio predak današnjih ptica, već samo “rođak.”
Ali kada mi skupljamo fosile predaka ptica, ova situacija predstavlja problem. Pogledajmo vremenski tok:
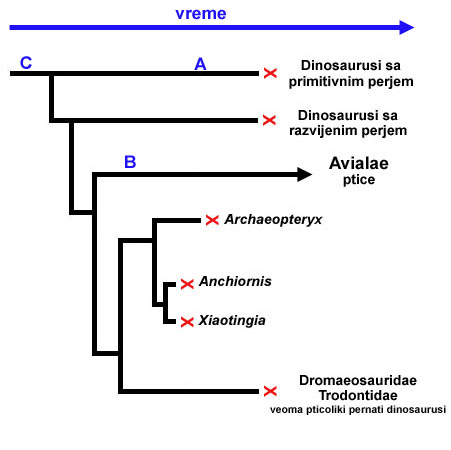
Mi možemo da nađemo fosil dinosaurusa sa primitivnim perjem iz tačke A. Zatim, možemo da nađemo fosil rane ptice na tački B. Na prvi pogled, može izgledati kao da je dinosaurus sa primitivnim perjem, predak ptica, mlađi od svog pticolikog potomka! Naravno, ovo je iluzija: poređenjem detalja telesne strukture fosila, mi možemo jasno videti gde se koji uklapa u ugnježdenu hijerarhiju.
Nezavisnu proveru možemo postići kroz nalaženje još fosila kojima možemo popuniti sliku, i kroz čije razlike možemo pratiti kako su se dešavale promene u morfologiji. Trenutno, naše poznavanje prelaza od dinosaurusa do ptica je zasnovano na više stotina fosila koji predstavljaju nekoliko desetina različitih vrsta.
Koliko su stabilne naše interpretacije fosilnih dokaza?
Sve naučne teorije se neprestano razvijaju u skladu sa novim dokazima i činjenicama obzerviranim u prirodi. Dobar primer ovoga je teorija gravitacije.
U sedamnaestom veku, Isak Njutn je razvio svoju teoriju gravitacije, zasnovanu na merenjima, podacima i obzervacijama koje su u njegovom dobu ljudima bile dostupne. Njegova teorija je dala matematičku osnovu razumevanja gravitacije, od kretanja granate ispaljene iz topa, do kretanja planeta oko Sunca. Ali ubrzo su nađene neke situacije kod kojih Njutnove jednačine nisu davale tačno rešenje: recimo, orbita Merkura se nije uklapala u Njutnove rezultate. Nalazi iz prirode su ukazali da teorija nije savršena, i da je potrebno proizvesti bolju.
Rešenje je našao Ajnštajn, koji je otkrio teoriju generalnog relativiteta, i dao neuporedivo bolju i jasniju sliku gravitacije. Pomoću Ajnštajnovih jednačina, moguće je rešiti probleme kao što je orbita Merkura, i dobiti tačna predviđanja za mnoge probleme kod kojih Njutnove jednačine daju netačna rešenja.
No, Ajnštajnovo otkriće ne znači da je Njutnova teorija netačna. Fizičari mogu i dan danas da pomoću Njutnovih jednačina izračunaju putanju objekta u padu, da izgrade most, ili da lansiraju rakete. Rezultati Njutnovih računica koji su bili tačni u sedamnaestom veku nisu odjednom postali netačni kada je Ajnštajn objavio svoju novu teoriju. Ajnštajnova teorija daje jasniju sliku gravitacije nego Njutnova, omogućava da se više problema reši, i da se dobiju tačniji rezultati.
Isti princip važi i za našu interpretaciju fosila. Što više fosila imamo, što je bolji kvalitet tih fosila, utoliko je bolje i naše razumevanje fosilnih prelaza. Zaključci koji se prave na osnovu nekolicine fosila su veoma nesigurni, i postaju utoliko sigurniji ukoliko se više fosila nakupi.
Možemo se ovde vratiti na prethodni primer fosila predaka ptica, i pogledati kako se naše razumevanje razvijalo. Naime, prvi pravi prelazni fosil, Archaeopteryx, je nađen još u devetnaestom veku, i skoro sto godina je bio jedini poznati fosil ptičjih predaka. Naše razumevanje celog ovog prelaza je u tom periodu izgledalo otprilike ovako:
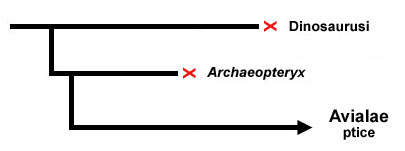
Imali smo osnovnu ideju da se od dinosaurusa (ili, moguće, srodnih arhosaurusa) odvojila grupa vrsta, među kojima su bili i Arheopteriks, i preci modernih ptica. Pošto je Arheopteriks bio jedina poznata vrsta (iz nekoliko različitih fosila), naučnici su formirali hipoteze o nastanku ptica i o nastanku perja samo na osnovu njega.
Ali krajem dvadesetog i početkom dvadeset prvog veka, ova slika se potpuno promenila: nađeni su mnogobrojni fosili, desetine vrsta pernatih dinosaurusa (dakle, dinosaurusa koji nisu ni ptice ni preci ptica), kao i fosili desetina vrsta ranih ptica. Ovo je omogućilo naučnicima da izgrade daleko jasniju sliku o ovom prelazu. Ta slika je ekstremno kompleksna, ali uprošćena verzija bi izgleda otprilike ovako:
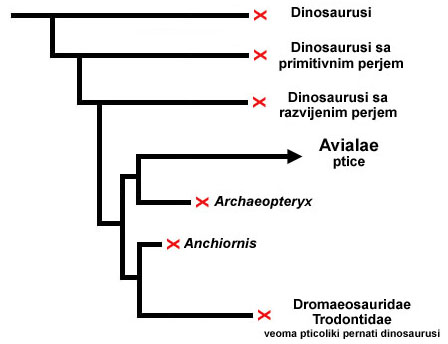
Drugim rečima, sada smo mogli da direktno na fosilima vidimo da su perje i ptičja struktura pluća nastali kod dinosaurusa daleko pre prvih ptica. Od veoma malih pernatih dinosaurusa (otprilike veličine prosečne kokoške), nastala je grupa srodnih vrsta veoma sličnih pticama, među kojima je bio i Archaeopteryx.
Zatim, 2011-te godine, nedugo pre pisanja ovog teksta, otkriće fosila pticolikog dinosaurusa Xiaotingia je donelo još podataka. Novo otkriće je omogućilo bolju analizu podataka, i ukazalo je na mogućnost da je Archaeopteryx bio bliži srodnik pticolikim dinosaurusima (kao što su Dromeosaurusi), nego ranim dinosaurolikim pticama. Ako je ova interpretacija tačna, shema nasleđa bi izgledala ovako:
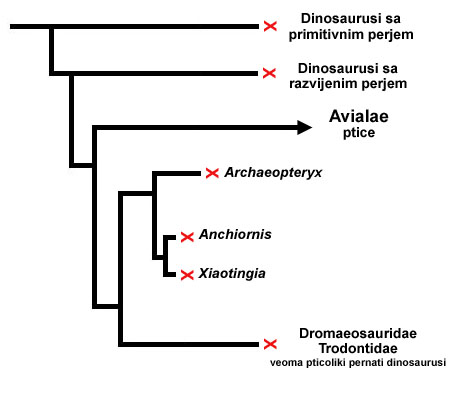
Sad, koja je tačna shema nasleđa? Dok ne nađemo još fosila iz ovog prelaza, ne znamo. Ali na sreću, to nije od kritične važnosti. Bez obzira da li je Arheopteriks “stric” današnjih ptica, ili je “brat dede,” on i dalje predstavlja vrstu koja je morfološki tek preko pola puta između pernatih dinosaurusa i pravih ptica. On je primer prelaznog fosila: vrste koja nam omogućava da vidimo kako se evolucija odvijala u prošlosti, i kako su se osobine menjale kroz vreme.
Napomena: označavanje “dubokog vremena”
Kada su naučnici počeli da razotkrivaju ogromnu starost sveta i nalaze dokaze da je život postojao mnogo duže nego što je iko ikada ranije i sanjao, pojavio se jedan novi jezički problem: naučni tekstovi su svakih pet reči pominjali milione i milijarde godina, toliko učestalo da je čitanje tekstova postajalo problem.
Zbog ovog, razvijena je konvencija označavanja starosti fosila i stena, zasnovana na metričkom sistemu. Konvencija je veoma jednostavna:
– starost u hiljadama godina se označava slovima Ka
– starost u milionima godina se označava slovima Ma
– starost u milijardama godina se označava slovima Ba
Dakle, ako se za nešto tvrdi da je staro deset hiljada godina, može se reći “to je staro 10Ka.” Određeni fosil, recimo stromatolit, star 3200 miliona godina, može biti označen kao fosil starosti 3200Ma, ili 3.2Ba. Radi jednostavnosti, ovu konvenciju ćemo koristiti u ostatku ovog teksta.
Nakon ovog uvoda, možemo da počnemo da pratimo fosile koji nam pokazuju kako se život na Zemlji razvijao. U sledećem poglavlju, pozabavićemo se najstarijim živim bićima, jednoćelijskim organizmima od kojih je sve počelo…