
Sadržaj:
- Uvod – šta su to prelazni fosili
- Razvoj višećelijskih organizama i Kambrijska eksplozija
- Razvoj kičmenjaka
- Od riba do vodozemaca
- Od vodozemaca do reptila
- Od reptila do sisara
- Od reptila do ptica
- Kratak pregled evolucije biljaka
- Žirafe, kitovi, konji i druge napomene
8. Kratak pregled evolucije biljaka
Implikacije
Evolucija biljaka je makar isto onoliko kompleksna koliko i evolucija životinja, ako ne i kompleksnija. Ovde ćemo dati samo najopštiji pregled kada nalazimo koje tipove biljaka u fosilnom zapisu, da bi smo pokazali još jedan značajan dokaz evolucije.
Naime, razvoj biljaka je u mnogim tačkama paralelan razvoju životinja; moderne biljke sa semenjem, recimo, su počele da se razvijaju u toku perioda Jure. Moderne trave su se razvile tek nakon nestanka dinosaurusa. Nikada, nigde nije nađen ni jedan jedini fosil dinosaurusa u istom sloju sa ostacima trave, ili u sloju iznad takvih ostataka. Nikada nisu nađeni ostaci trave u gnezdima dinosaurusa, niti u njihovim stomacima. Nikada, nigde nije naden ni jedan fosil ničega, uopšte, zajedno sa fosilima biljaka koje ne spadaju u isti vremenski period.
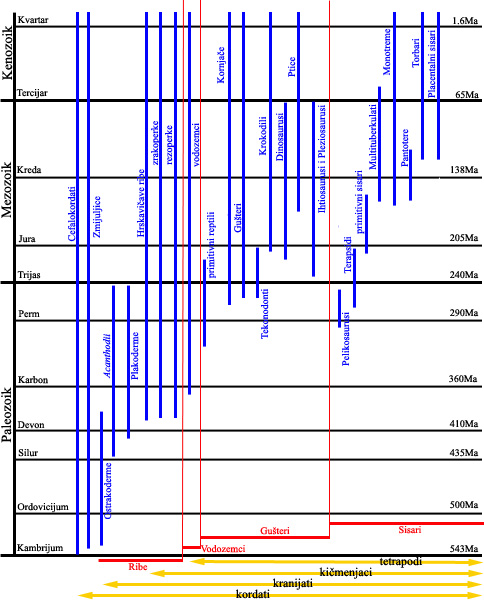
Pogledajmo paralelno veoma uprošćenu shemu razvoja kordata:
I jednu još uprošćeniju shemu razvoja biljaka:
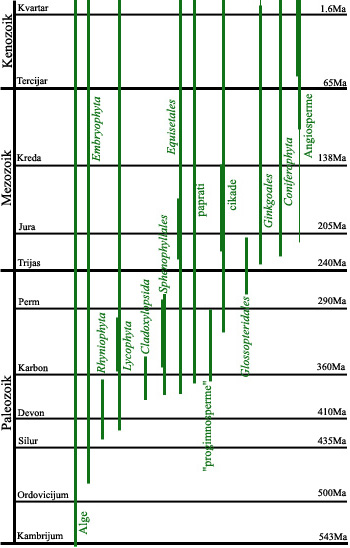
Pomoću ove dve sheme može da se vidi generalna slika koje životinje i koje biljke su postojale istovremeno u određenom periodu razvoja života na Zemlji. Imajte u vidu da ove sheme prikazuju samo velike, opšte grupe: podgrupe i pogrupe podgrupa i životinja i biljaka su se odvjale od ovih osnovnih linija u raznim periodima…i to odvajanje možemo da pratimo ne samo pomoću radiometrijskog datiranja i pomoću praćenja slojeva zemljišta u kojima su uzorci nađeni, već i kroz usklađivanje životinjskih i biljnih vrsta nađenih zajedno.
Izlazak biljaka na kopno
Kao što smo videli u prvom poglavlju ovog teksta, alge su među prvima razvile višećelijske organizme, koji su već imali primitivno stablo (sastavljeno samo od par jedva diferenciranih vrsta ćelija) i rizome, primitivne strukture koje su služile da ih drže pričvršćenim za tlo. Tri glavne prepreke stoje između prelaza tih prvobitnih algi na suvo tlo.
Prvi problem je reprodukcija. Kao i većina drugih eukariota, alge poseduju sposobnost seksualne reprodukcije, u kojoj se jedna ćelija sa dve kopije svakog hromozoma (takozvana diploidna ćelija) razdvaja na dve ćelije sa po jednom kopijom svakog hromozoma (takozvana haploidna ćelija). U vodi, proces reprodukcije je jednostavan: dve haploidne ćelije plivaju kroz vodu, nađu jedna drugu, spoje se u diploidnu ćeliju, i izrastu u novi organizam. Na suvom, ovaj proces je mnogo komplikovaniji.
Drugi problem je tečnost. Alga izbačena na suvo tlo će se ubrzo osušiti, pošto ne poseduje mehanizme koji bi joj omogućili da zadrži vodu unutar stabla.
 Treći problem je prenos hranjivih sastojaka. Alge u vodi mogu da upijaju minerale direktno iz vode koja ih okružuje. Na suvom, minerali iz zemljišta moraju nekako biti preneseni do drugih delova biljke.
Treći problem je prenos hranjivih sastojaka. Alge u vodi mogu da upijaju minerale direktno iz vode koja ih okružuje. Na suvom, minerali iz zemljišta moraju nekako biti preneseni do drugih delova biljke.
Razvoj kopnenih biljaka počinje sa razvojem grupe poznate kao Embryophyta, koja se razvila iz veoma sličnih vodenih Chlorophyta, zelenih algi. Prve embriopite su bile biljke koje su rasle samo na veoma vlažnom i stalno kvašenom zemljištu, i bile su visoke samo nekoliko milimetara do maksimalno par centimetara. Ovim su sva tri problema izbegnuta: haploidne ćelije (spermatozoidi) i dalje mogu da plivaju kroz vodu do svog cilja, voda stalno kvasi biljku i donosi hranjive sastojke, a njihov prenos se obavlja kroz statički pritisak vode unutar biljke.
Ali ove biljke su bile veoma osetljive – povlačenje vode je značilo sušenje i smrt. Ovo je ubrzo dovelo do razvoja epiderma, voštanog sloja koji prekriva biljku i sprečava njeno isušivanje. Epiderm predstavlja napredak, ali i problem: prilikom suvog perioda, pošto voda ne može da isparava slobodno iz biljke, ona se na suncu može lako pregrejati. Zbog ovoga, rane embriofite razvijaju drugu ključnu osobinu: stome, parova ćelija koje proizvode kontrolisane pore u lišću, i omogućavaju da voda ipak isparava dovoljno da bi se temperatura mogla kontrolisati.
Rizomi nekih embriofita, osim svoje osnovne funkcije da drže biljku vezanom za jedno mesto, takođe počinju da upijaju minerale iz okolnog zemljišta. Ove strukture su, međutim, i dalje daleko od prvih pravih korena.
 Sledeci korak osvajanja kopna vidimo kod biljaka koje su pripadale danas izumrloj grupi poznatoj kao Rhyniophyta, u koju spada i biljka poznata kao Cooksonia pertoni (fosil na slici). Arkegonium, organ koji drži “ženske” haploidne ćelije, ostaje vezan za biljku, umesto da bude prosto izložen na površini. Stablo postaje izduženo i dihotomno se grana (jedna grana se razdvaja na dve manje, još uvek nema specijalizovanog lišća), sa muškim seksualnim organom, sporangijom, na krajevima.
Sledeci korak osvajanja kopna vidimo kod biljaka koje su pripadale danas izumrloj grupi poznatoj kao Rhyniophyta, u koju spada i biljka poznata kao Cooksonia pertoni (fosil na slici). Arkegonium, organ koji drži “ženske” haploidne ćelije, ostaje vezan za biljku, umesto da bude prosto izložen na površini. Stablo postaje izduženo i dihotomno se grana (jedna grana se razdvaja na dve manje, još uvek nema specijalizovanog lišća), sa muškim seksualnim organom, sporangijom, na krajevima.
Najprostije riniofite (kao što je, recimo, Horneophyton) nemaju nikakav sistem prenosa tečnosti, ali kasnije forme pokazuju prvi razvoj traheida, kanala koji prenose tečnost kroz stablo biljke. Ovi kanali su omogućili biljci da se manje oslanja na pritisak vode, i da više izraste: najveće riniofite su bile i do 20cm visoke. Riniofite su davno izumrle, ali se generalno smatraju direktnim precima svih kopnenih biljaka osim mahovine.
Kasne riniofite, i još neke prelazne grupe iz ovog perioda, (zosterofilofite, na primer) pokazuju dalji razvoj transportnih tkiva do prve diferencijacije između ksilema i floema. Ksilem je prosta serija kanala koji kapilarnim pritiskom vuku vodu iz donjih delova biljke u gornje. Floem je slična serija kanala, koja prenosi šećere proizvedene fotosintezom i minerale u sve delove biljke pomoću osmotskog pritiska.
Lycophyta ssu ovu osnovnu strukturu dovele do maksimuma. Ova grupa biljaka je razvila strukture slične lišću, takozvane mikrofile, koji se sastoje od jednog vaskularnog kanala za prenos tečnosti koji raste van stabla biljke. Mada ni približno onoliko efikasne kao moderno lišće, ovaj sistem je značajno unapredio sposobnost biljke za fotosintezu, i doveo do ogromne eksplozije biljnog života na kopnu u doba Karbona (360-290Ma).
Likofite su počele da proizvode materijale slične ligninu, i da izrastaju u sve čvršća i čvršća stabla. Na vrhuncu razvoja, ogromna stabla ovih biljaka su bila i po 35 metara visoka, i (zajedno sa potomcima likofita o kojima će biti reči u daljem tekstu) prekrivala su kopno ogromnim šumama. Geohemijska merenja pokazuju da je količina ugljen-dioksida u atmosferi u ovom periodu opala sa 3600ppm na oko 300ppm u eksploziji fotosinteze i reakcije silikatnih stena sa organskim kiselinama koje je oslobađao raspad biljnog tkiva. Novi drvenasti materijali su takođe bili nešto novo – jednoćelijski organizmi nisu još bili prilagođeni za njihovo brzo razlaganje. Ovo je dovelo do akumulacije organskog materijala tokom miliona godina, koji je bio prekriven daljim sedimentnim i vulkanskim slojevima, i konačno se pretvorio u ugalj.
Sa razvojem transportnih sistema i drvenastog stabla koje može da podnese veliku visinu i težinu biljke, rešeni su svi glavni problemi osim reproduktivnih. Biljke iz perioda Karbona su se i dalje razmnožavale pomoću spora, koje su zahtevale veliku količinu vlage. Ali i pored toga, biljke su se našle u poziciji u kojoj su im ogromne površine Zemlje postale otvorene za naseljavanje. Naglo širenje je dovelo do nagle radijacije biljaka na ogroman broj različitih osnovnih vrsta; u određenom smislu, kraj Devonskog perioda je period eksplozije kopnenog biljnog života na sličan način kao što je broj životinjskih vrsta eksplodirao u Kambrijskom periodu nekih dve stotine miliona godina ranije.

(Na slici je rekonstrukcija izgleda šume iz perioda Karbona: u sredini je likofita poznata kao Sigillaria; možete primetiti da nema grana, već da lišće raste direktno iz stabla. U pozadini sa leve strane je velika paprat, a sa desne stablo iz porodice Equisetopsida.)
Equisetopsida je jedna od grupa koje su nastale u ovom periodu. Veoma slične po građi likofitama, ekvisetopside su razvile mikrofile u prvo pravo lišće – mada je ono još uvek bilo veoma primitivno u poređenju sa lišćem modernih biljaka. Ove biljke su se u periodu Karbona razvile na više strana; neke su postale ogromne, sa stablima visokim i do 30m, dok su druge rasle u obliku žbunja. Početkom Perma, dolazi do masovnog istrebljenja u kome nestaje preko 90% životinjskih vrsta na zemlji, a zajedno sa njima umiru i ogromne Karbonske šume. Ekvisetopside će opet postati značajne tokom kratkog perioda u toku Trije, ali će zatim nastaviti da odumiru. Danas je od cele ove ogromne grupe biljaka preživelo samo dvadesetak vrsta iz jednog roda, Equisetum, u narodu poznat kao “konjski rep”.
 Pteridopsida, grupa koja uključuje paprati i još nekoliko izumrlih grupa biljaka, su bile bolje sreće. Ogromne paprati, i do deset metara visoke, su vladale šumama tokom doba dinosaurusa, a značajan broj vrsta je preživeo sve do danas, prevashodno u kišnim šumama. Paprati su razvile prosto lišće kakvo vidimo kod ekvisetopsida u karakteristične široke multitubularne izraštaje koje danas prepoznajemo. Korenje, koje je počelo da se razvija u glavni absorptivni organ još kod ranih likofita, kod paprati postaje potpuno razvijeno. Vaskularni sistem i slojevita struktura lišća takođe postaju daleko razvijenije nego bilo šta viđeno kod prethodnih vrsta.
Pteridopsida, grupa koja uključuje paprati i još nekoliko izumrlih grupa biljaka, su bile bolje sreće. Ogromne paprati, i do deset metara visoke, su vladale šumama tokom doba dinosaurusa, a značajan broj vrsta je preživeo sve do danas, prevashodno u kišnim šumama. Paprati su razvile prosto lišće kakvo vidimo kod ekvisetopsida u karakteristične široke multitubularne izraštaje koje danas prepoznajemo. Korenje, koje je počelo da se razvija u glavni absorptivni organ još kod ranih likofita, kod paprati postaje potpuno razvijeno. Vaskularni sistem i slojevita struktura lišća takođe postaju daleko razvijenije nego bilo šta viđeno kod prethodnih vrsta.
Progimnosperme su ključna vrsta iz ovog perioda, pošto su one najverovatniji preci današnjih biljaka sa semenom. Kod ove grupe se prvi put pojavljuje pravo drvo, koje je sačinjeno od sekundarnih slojeva vaskularnog tkiva. Drugim rečima, kanali koji prenose hranjive materije i vodu rastu i razvijaju se u mnogo debljim slojevima nego kod drugih biljaka, i kada se konačno sasuše, postaju strukturalna podrška za težinu biljke. Takođe, struktura ksilema poseduje perforacije koje su veoma slične onima koje vidimo kod četinara.
Ove biljke su se, međutim, i dalje razmnožavale pomoću spora, i njihovo lišće je po građi bilo veoma slično papratima.
Spermatophyta, biljke sa semenom
Spermatophyta su grupa biljaka sa kojima je današnji čovek najbolje upoznat. Ova grupa danas obuhvata ogroman broj vrsta, od trave do hrastova, ali je u prvom delu svoje istorije igrala relativno malu ulogu u opštoj slici zemaljske flore.
Najstarije biljke sa semenom su vrste paprati, takozvane “semene paprati”; najstariji poznat fosil je Elkinsia polymorpha. Ova grupa biljaka je razvila primitivno seme (bez cvetova, šišarki, ili bilo kakvih drugih kompleksnih struktura) prostom modifikacijom lišća: list bi izrastao oko semena, osušio se i očvrsnuo. Za razliku od spora, seme je otpornije na promene u temperaturi i vlažnosti, zaštićenije je od mikroorganizama i drugih oblika života koji mogu da ga napadnu, i može da odloži period rasta do trenutka kada su uslovi pogodni.
Na vrhu ovog primitivnog semena se nalazio otvor, mikropil, kroz koji je “muška” spora prolazila da bi oplodila seme. Paralelno sa razvojem strukture semena, struktura spora se menjala i postajala sve sličnija polenu. Kod kasnijih spermatofita, odmah ispod mikropila se razvija komora obložena lepljivom tečnošću koja pomaže da se polen “zarobi” – prvi početak današnjeg nektara.
 Semene paprati su potpuno izumrle do danas, kao i nekoliko drugih grupa grupa spermatofita iz ovog ranog perioda (npr. Medullosales). Međutim, njihovi potomci će postati najuspešnije biljke u istoriji života na Zemlji.
Semene paprati su potpuno izumrle do danas, kao i nekoliko drugih grupa grupa spermatofita iz ovog ranog perioda (npr. Medullosales). Međutim, njihovi potomci će postati najuspešnije biljke u istoriji života na Zemlji.
Cycadophyta, ili cikade, su prva klasa potpuno razvijenih spermatofita nazvanih gimnosperme (“golo seme”). Sa mnogim osobinama paprati (svo lišće raste iz jednog centra), ali i sa semenjem, ove biljke su se naglo razvile u toku perioda Trije, kada je klima postala mnogo suvlja i toplija, i u velikoj potisnula biljke koje su zavisile od spora. U toku perioda Jure, oko jedna petina svog kopnenog biljnog života su bile cikade. Mada je većina vrsta izumrla u toku perioda Krede, mnoge vrste su opstale i do danas.
 Glossopteridae su misteriozna grupa biljaka koja je bila prisutna u toku zadnjeg dela Perma i ranog dela Trije – misteriozna zato što do dana današnjeg niko nije siguran kako su biljke iz ove grupe izgledale. Iz nekog razloga, njihova stabla se nisu fosilizovala u celini, već u sitnim komadima koje niko ne zna kako da uklopi u celinu. Dve stvari čine glosopteride zanimljivim za našu analizu. Prvo, njihovo lišće je pokazuje liniju kidanja, pomoću koje se lišće listopadnog drveća odvaja od stabla. Mnogo važniji, međutim, je njihov način razmnožavanja. Muški organi sa polenom su se nalazili na zadnjoj strani lišća, slično kao kod paprati; ali ženski organi su kod nekih vrsta bili grupisani unutar posebne vrste lišća koja se savijala oko njih čineći primitivan cvet.
Glossopteridae su misteriozna grupa biljaka koja je bila prisutna u toku zadnjeg dela Perma i ranog dela Trije – misteriozna zato što do dana današnjeg niko nije siguran kako su biljke iz ove grupe izgledale. Iz nekog razloga, njihova stabla se nisu fosilizovala u celini, već u sitnim komadima koje niko ne zna kako da uklopi u celinu. Dve stvari čine glosopteride zanimljivim za našu analizu. Prvo, njihovo lišće je pokazuje liniju kidanja, pomoću koje se lišće listopadnog drveća odvaja od stabla. Mnogo važniji, međutim, je njihov način razmnožavanja. Muški organi sa polenom su se nalazili na zadnjoj strani lišća, slično kao kod paprati; ali ženski organi su kod nekih vrsta bili grupisani unutar posebne vrste lišća koja se savijala oko njih čineći primitivan cvet.
Sledeća značajna grupa su Ginkgophyte, koje su se odvojile od cikadolikih predaka i razvile šire lišće i mesnatije seme. Ginkofite pokazuju prve znake zaštite semena od životinja: prilikom truljenja, seme oslobađa veliku količinu butirne kiseline, što ga čini veoma smrdljivim, a mesnati deo sadrži značajnu količinu cijanida. Takođe, potpuno je sigurno da se radi o listopadnom drveću.
Ova grupa biljaka je bila veoma uspešna tokom Mezozoika, sa šesnaest poznatih rodova i ogromnim brojem različitih vrsta. Najveći broj njih je, međutim, nestao zajedno sa dinosaurusima, a preostale vrste su se polako gasile jedna za drugom tokom Kenozoika. Rani naučnici su dugo smatrali da je ova čitava klasa potpuno izumrla, i bili su skoro u pravu – od šuma ginkofita koje su prekrivale kontinente, od desetina različitih vrsta, preživelo je samo sedam stabala vrste Ginkgo biloba. Tih sedam stabala je otkriveno krajem osamnaestog veka u jednom budističkom manastiru u istočnoj Kini, i njihovo seme je odmah rašireno širom sveta. Danas je Ginkgo biloba jedno od najčešce gajenih ukrasnih stabala na svetu, sa raznim medicinskim primenama.
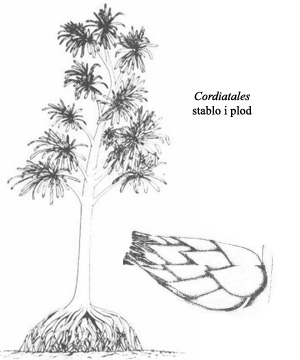 Grupa sporofita poznata kao Cordiatales je davno izumrla, ali je važna za našu diskusiju pošto predstavlja prelazni oblik u razvoju četinara. Lišće je i dalje veliko, ali postaje izduženo, sa vaskularnom strukturom sličnom današnjim četinarima. Razvoj ploda se odigrava u posebnim lisnatim izraštajima: posebno lišće raste tako da se preklapa jedno sa drugim, dok se plod nalazi između lisnatih slojeva. Ovo lišće se verovatno sušilo i postajalo čvrsta zaštita za plod, iz koga ga životinje nisu mogle lako izvući (ptice još nisu postojale, tako da nije bilo ni kljunova koji su mogli da iščupaju plod iz takve ljušture).
Grupa sporofita poznata kao Cordiatales je davno izumrla, ali je važna za našu diskusiju pošto predstavlja prelazni oblik u razvoju četinara. Lišće je i dalje veliko, ali postaje izduženo, sa vaskularnom strukturom sličnom današnjim četinarima. Razvoj ploda se odigrava u posebnim lisnatim izraštajima: posebno lišće raste tako da se preklapa jedno sa drugim, dok se plod nalazi između lisnatih slojeva. Ovo lišće se verovatno sušilo i postajalo čvrsta zaštita za plod, iz koga ga životinje nisu mogle lako izvući (ptice još nisu postojale, tako da nije bilo ni kljunova koji su mogli da iščupaju plod iz takve ljušture).
Kod coniferophyta, četinara, lišće postaje manje i gušće, sa češćim grananjem. Lisnata ljuštura se razvija i očvršćuje, dok kod kasnijih, razvijenih četinara ne postane ono što danas nazivamo šišarka (mnogi primitivniji četinari i dalje imaju ljušturu koja je na pola puta između lisnate ljuske Cordiatalesa i modernih šišarki). Slojevi voska oko epiderma postaju deblji, a površina lišća se smanjuje. Unutrašnje tečnosti postau bogate smolom, koja deluje kao antifriz i omogućava drveću da lakše preživi u veoma hladnim krajevima. četinari su na osnovu ovih adaptacija zauzeli ogromne hladne oblasti i formirali tajge kakve vidimo i danas.
Gnetophyta je rod biljaka koji pokazuje neobičnu mešavinu osobina gimnospermi i angiospermi (sledeće poglavlje), što je dovelo do nekih teorija da su biljke iz ovog roda prelazni oblik između ove dve grupe. Međutim, moderne genetske analize pokazuju veću srodnost sa četinarima nego sa cvetnim biljkama, što je potvrđeno novijim fosilnim nalazima koji početak angiospermi stavljaju u doba Trije. Po ovoj novijoj interpretaciji, Gnetofite su se odvojile od predaka angiospermi na samom početku mezozoika. Implikacija ovoga je da su mnoge osobine današnjih angiospermi, uključujući tu i dvostruku oplodnju (jedno zrno polena oslobađa dva spermatozoida, jedan da oplodi jaje, drugi da izraste u endosperm, izvor energije za rast semena), nastale veoma rano u razvoju biljaka.
 Nekoliko vrsta gnetofita je preživelo do danas. Najpoznatiji predstavnici su Welwitschia mirabilis, biljka sa najvećim lišćem na svetu, i efedra, poznata po svom ekstraktu koji se koristi za povećanje metabolizma i mršavljenje (ovaj ekstrakt je postao zloglasan i zabranjen na nekim tržištima, pošto može da izazove srčane udare).
Nekoliko vrsta gnetofita je preživelo do danas. Najpoznatiji predstavnici su Welwitschia mirabilis, biljka sa najvećim lišćem na svetu, i efedra, poznata po svom ekstraktu koji se koristi za povećanje metabolizma i mršavljenje (ovaj ekstrakt je postao zloglasan i zabranjen na nekim tržištima, pošto može da izazove srčane udare).
Angiosperme
Angiosperme su dugo bile misterija sa stanovišta fosilnog zapisa; najranije vrste iz ove grupe nisu bile drvenaste, i veoma retko su se fosilizovale. Međutim, u zadnjih dvadesetak godina je nađeno nekoliko fosila vrsta koje pokazuju mnoge primitivne osobine angiospermi, tako da slika polako postaje sve jasnija. Molekularne analize takođe pomažu da se razluči stablo razvoja angiospermi.
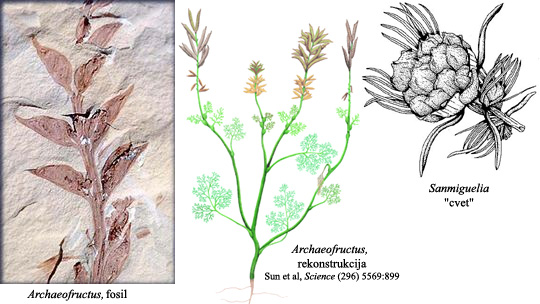
Sanmiguelia je ključni fosil u ovom prelazu. Ovaj fosil iz kasnog perioda Trije pokazuje mnoge osobine kasnijih angiospermi, uključujući dvostruku oplodnju, organizovani kambijum u centru stabla, i sa jasno razvijenim sekundarnim rastom. Što je još značajnije, Sanmiguelia pokazuje mešavinu osobina dikotila i monokotila, dve osnovne klase angiospermi. Cvetolike strukture su veoma slične šišarkama angiospermi, jedva nešto više od lišća koje raste u povezanom obruču; ali svi potrebni elementi za buduće cvetove su prisutni.
Archaeofructis je biljka iz perioda Jure sa daljim razvojem ka modernim cvetnim biljkama. Organizacija organa za razmnožavanje odgovara cveću, i polen je na izduženim stamenima. Niz drugih osobina koje se pojavljuju kod kasnijih angiospermi je takođe prisutan, ali su prisutne i razlike koje sprečavaju da se Archaeofructis smatra prvom pravom angiospermom.
U periodu Krede dolazi do eksplozije različitih vrsta angiospermi, u razmerama koje daleko prevazilaze okvire ovog teksta. Verovatan razlog za ovo je koevolucija insekata. Insekti su počeli da napadaju i jedu plodove angiospermi od samog početka: najbolji fosil Sanmiguelie ima jasno vidljive tragove ujeda insekta blizu osnove ploda. Mada su angiosperme još dugo vremena nastavile da se oslanjaju prevashodno na vetar za prenos polena, insekti su postajali sve važniji. Najverovatniji razlog za naglu eksploziju angiospermi u toku Krede je upravo ova koevolucija, ili konkretnije, razvoj slatkog nektara sa svrhom da mami insekte ka polenu.
Desetine fosila iz ovog perioda pokazuju rani razvoj cveća (razlika između monokotila i dikotila je već ustanovljena do ove tačke, i poznata je samo iz implikacija mešavine osobina kod Sanmiguelie i iz molekularnih analiza), ali mnoge primitivne četinarske osobine su opstale sve do danas. Konkretno, magnolije, koje su među najprimitivnijim cvetnicama, imaju veoma primitivan cvet i plod koji je veoma sličan šišarkama (ne samo po spoljnom izgledu, već i po unutrašnjoj građi i po genetskoj stukturi).

[prethodno poglavlje][nazad na vrh strane][sledeće poglavlje]