
- Uvod – šta su to prelazni fosili
- Razvoj višećelijskih organizama i Kambrijska eksplozija
- Razvoj kičmenjaka
- Od riba do vodozemaca
- Od vodozemaca do reptila
- Od reptila do sisara
- Od reptila do ptica
- Kratak pregled evolucije biljaka
- Žirafe, kitovi, konji i druge napomene
4. Od riba do vodozemaca
U prethodnom poglavlju smo obradili evoluciju kičmenjaka i riba, do tačke na kojoj su se razdvojile zrakoperke i rezoperke. Pogledajmo sada kako je tekao prelaz od riba rezoperki do vodozemaca.
U okviru nastanka kopnenih životinja, prvi ključni korak se odigrao daleko pre nego što je bilo koja životinja stupila na suvo tlo. Molekularni, genetski, i morfološki dokazi pokazuju da su već rane koščate ribe posedovale takozvanu gastričku kesicu: proširenje sa strane jednjaka, u koje je riba mogla da “proguta” vazduh, što olakšava plivanje (ribe sa ovakvim kesicama mogu da plutaju, dok ribe bez njih – na primer, ajkule – moraju neprestano da ulažu energiju i plivaju, jer u suprotnom potonu na dno okeana). Ova gastrička kesica je postala predmet selekcije. Kod riba u otvorenoj vodi, ona je postala ključna za plivanje, i razvila se u današnju vazdušnu bešiku.
Dodatni podaci o prelaznim fosilima
Ovaj tekst se fokusira na ključne fosile. Postoji mnogo više prelaznih oblika, čije opise možete naći da na mestima kao što je Palaeos.
Kod riba u plitkim i mutnim vodama siromašnim vazduhom, ova kesica se nalazila pod drugačijim selektivnim pritiskom. U ovakvoj vodi ribi je dostupno veoma malo kiseonika, a blato koje zaustavlja protok kroz škrge čini stvari još gorim. Otud, gastrička vrećica je kod njih postala prokrvljena, omogućavajući ribi da iz progutanog vazduha apsorbuje kiseonik. Takođe, svaka mutacija koja pomera otvor kesice bliže ustima je pozitivno selektovana, pošto smanjuje verovatnoću da progutani vazduh završi u želucu (gde kiseonik ne može efikasno da se apsorbuje).
Ovaj put razvoja je doveo do nastanka primitivnih ribljih pluća, koja možemo i dan danas da vidimo kod nekoliko ribljih grupa. Primer ovoga su ribe gar i amija među zrakoperkama, i riba dvodihalica među rezoperkama.

Kvinslendska dvodihalica, Neoceratodus forsteri, poseduje škrge, pluća i koščata peraja.Photo: Tannin
Pre oko 400 miliona godina, kopnene biljke su konačno počele da stabilizuju priobalne ekosisteme, što je vodenim životinjama pružilo mogućnost novih načina života, i novih izvora hrane. Usled suše, ili čak prosto ciklusa plime i oseke, ove oblasti su često ostajale izolovane (u vidu muljevitih bara odvojenih od otvorene vode) i bile su siromašne kiseonikom. Otud, nije čudno što u fosilnim nalazištima iz priobalnih ekosistema ovog perioda često nalazimo upravo mnogo riba dvodihalica. Jedna grupa ovakvih riba, po imenu ripidistija (Rhipidistia), je posebno značajna.
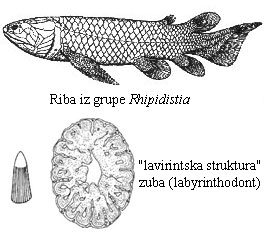 Ove ribe, kao i sve dvodihalice, su imale i pluća i škrge. Takođe, dok su “zraci” u perajima zrakoperki vezani direktno za osnovni skelet, kod ripidistijanskih riba i drugih rezoperki, ovi “zraci” su vezani za snažne kosti i mišiće. Konačno, imali su i osobinu koja je veoma jedinstvena za ovu grupu: njihovi zubi su imali veoma specifičnu “lavirintsku” strukturu.
Ove ribe, kao i sve dvodihalice, su imale i pluća i škrge. Takođe, dok su “zraci” u perajima zrakoperki vezani direktno za osnovni skelet, kod ripidistijanskih riba i drugih rezoperki, ovi “zraci” su vezani za snažne kosti i mišiće. Konačno, imali su i osobinu koja je veoma jedinstvena za ovu grupu: njihovi zubi su imali veoma specifičnu “lavirintsku” strukturu.
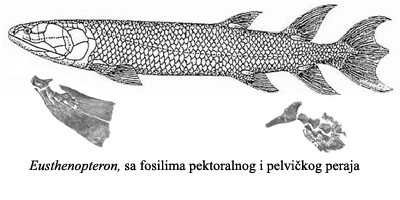
U fosilnom zapisu vidimo kako su se ripidistijanske ribe, izložene novim selektivnim pritiscima, brzo prilagodile svojoj novoj okolini. U okviru prilagođavanja na život u plićaku (umesto plivanja u dubokom moru), telo kasnijih ripidistija postaje spljošteno, oblik lobanje se menja, rep postaje prav. Zbog hroničnog nedostatka kiseonika, pluća postaju efikasnija i podeljena na dva dela. Konačno, kosti peraja postaju veće, snažnije, i mišići postaju snažniji. Ovakva peraja su skoro neupotrebljiva za plivanje kroz duboku vodu, ali su veoma korisna za razgrtanje gustog bilja u plićaku, i za puzanje kroz mulj. Primer jedne ovakve ribe je bio eustenopteron (Eusthenopteron):
Pritisci okoline su proizveli mnogo vrsta veoma bliskih Eusthenopteronu, ali koje su se još više odmakle od riba. Jeda lep primer ovoga je panderihtis (Panderichthys):

Autor: Nobu Tamura
Pošto više nije bilo selektivnog pritiska koji održava postojanje leđnih i analnih peraja, kod panderihtisa su ona zakržljala. Ostao je samo rep i kratka peraja na veoma snažnim i koščatim udovima. Povrh toga, kosti u okviru ovih udova su postale potpuno diferencirane, i to na veoma specifičan način: humerus, ulna i radius u prednjim udovima, i femur, tibija i fibula u zadnjim udovima. Takođe se pojavila čoana, prolaz u kosti koji povezuje nosnu šupljinu sa ustima, što omogućava disanje kroz nos.
Ovakve “puzeće vrste” se šire i granaju u toku sledećeg perioda vremena. Imamo mnogo prelaznih fosila iz ovog perioda, koji pokazuju razne vrste životinja sličnih panderihtisu (recimo Ventastega, ili Elginerpeton). Fosil koji nas posebno zanima je zvan tiktalik (Tiktaalik):
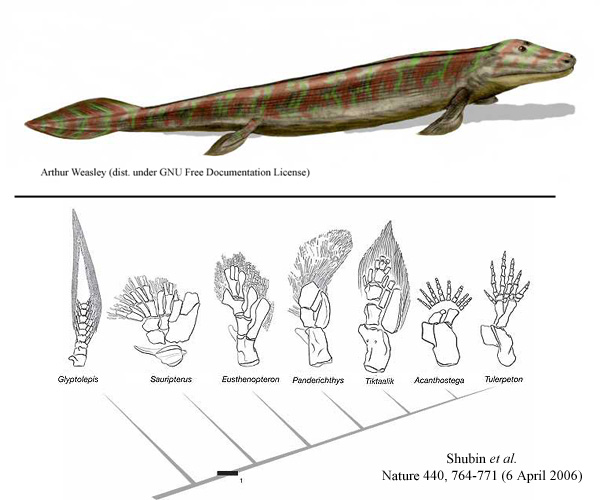
Kod tiktalika, vidimo još par malih promena (da ponovimo, svaki korak u evoluciji čine male, malecne promene koje se polako šire i nakupljaju). Najvažnije za nas su promene u strukturi udova: veza između kostiju koja je kod panderihtisa bila čvrsta, kod tiktalika postaje savitljiva (drugim rečima, nastaje zglob). Povrh toga, neki od “zraka” u okviru peraja postaju zadebljani, što omogućava životinji da se lakše odupre od tla, i samim tim da lakše puzi po zemlji; ova zadebljanja su u obliku niza falangijalnih kostiju (tj. kostiju prstiju).
Ova zadebljanja padaju pod selektivni pritisak za lakše kretanje kroz mulj i preko kratkih distanci na suvom tlu. Posle samo par miliona godina, vrste koje su nasledile ove osobine se šire svetom, toliko da neke od njih nalazimo u mnogim naslagama priobalnih područja u svetu. Jedan primer ovakve uspešne vrste je akantostega (Acanthostega):

Ova “žaboriba” je imala i pluća i škrge. Mada su kosti lobanje identične kasnijim vodozemačkim, oblik lobanje je još uvek riblji. Rep još uvek ima riblje karakteristike, sa perajem na leđnoj strani. Veza između lobanje i ostatka tela je riblja. Zubi i dalje imaju lavirintsku strukturu, nasleđenu od ripidistijanskih riba. I dalje postoji lateralna linija, organ koga ribe koriste da osete vibracije u vodi (ali koga vodozemci nemaju). Kada se sve uzme u obzir, nemoguće je reći da li se radi o ribi ili o vodozemcu – Acanthostega je savršena prelazna forma između dve klase živih bića.
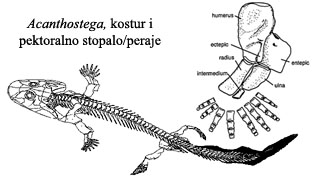
Kod akantostege, raspored i broj kostiju u “rukama” i “nogama” je identičan onome koga vidimo kod svih kopnenih kičmenjaka (guštera, ptica, sisara…). Pozicija stopala je, međutim, još uvek prilagođena vodenom životu, i nepogodna za hodanje po suvom tlu. Ovaj razvoj vidimo kod jedne srodne vrste iz istog perioda, ihtiostege (Ichtyostega):
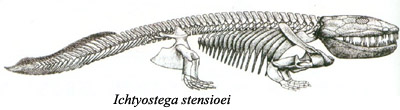
Udovi ihtiostege su prvi koji su direktno prilagođeni za hodanje po suvom tlu. Još uvek su prisutne razne riblje osobine: zglob između lobanje i kičme, oblik kičmenih kostiju, rep tipičan za rezoperke, lateralna linija… međutim, vodozemačke osobine su toliko izražene da se ihtiostega obično klasifikuje kao rani vodozemac.
Od ove tačke, rani vodozemci poput ihtiostege su se raširili svuda po svetu, i ubrzo se razdvojili na stotine raznih vrsta – Pholidogaster, Denderpeton, Tulerpeton (na donjoj slici)… . Neke od ovih vrsta su ostale vodozemci. Neke su čak ponovo napustile kopno, i vratile se potpuno vodenom životu (kao što je recimo bio slučaj sa krasigirinusom na donjoj slici; radi reference, ova životinja je bila preko metar i po dugačka, a njeni zakržljali udovi su bili dugi samo nekoliko centimetara). Evolucija nikada ne stoji, i adaptacije proizvode nove vrste u svim mogućim pravcima, sa svake moguće tačke…
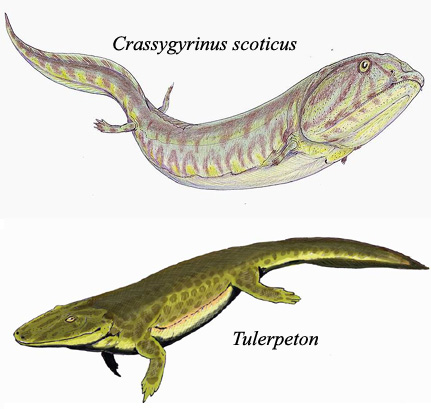
No, neke vrste su nastavile dalje da se razvijaju ka kopnenom životu. Jedna od ovih grupa, Anthracosauria, je postala predak ranim reptilima, kojima ćemo se pozabaviti detaljnije u sledećem poglavlju. Dva primera, Pholidogaster i Dendrerpeton su prikazana na slici.

[prethodno poglavlje][nazad na vrh strane][sledeće poglavlje]