
Sadržaj:
- Uvod – šta su to prelazni fosili
- Razvoj višećelijskih organizama i Kambrijska eksplozija
- Razvoj kičmenjaka
- Od riba do vodozemaca
- Od vodozemaca do reptila
- Od reptila do sisara
- Od reptila do ptica
- Kratak pregled evolucije biljaka
- Žirafe, kitovi, konji i druge napomene
6. Od reptila do sisara
Razvoj sisara od guštera je odlično dokumentovan u fosilnom zapisu. Kao što smo videli u prethodnom poglavlju, sinapsidi (klasa reptila od koje su nastali sisari) se odvojila od ostalih reptila veoma rano u istoriji njihovog razvitka. Evolucija sisara od sinapsida je od te tačke išla u dva koraka. Od ranih sinapsida je prvo evoluirala čitava klasa vrsta sa građom koja je otprilike na sredini između reptilske i sisarske: terapsidi. Zatim su od terapsida nastali prvi monotremni sisari, pa od njih sisari koje poznajemo danas.
Pre nego što počnemo sa pregledom fosilnog zapisa razvoja sisara, moramo napomenuti jednu stvar: kao što je već više puta rečeno, meka tkiva se jako retko fosilizuju, i na raspolaganju imamo uglavnom kosti. Iz ovog razloga, fosili nam ne mogu jasno pokazati kada su se prvi put razvile mlečne žlezde, ili krzno. Ali razvoj sisara možemo pratiti veoma detaljno na bitnih i sveobuhvatnih osnovu razlika u građi njihovog muskuloskeletalnog sistema. Katlin Hant, biolog sa Vašingtonskog Univerziteta je sklopila sledeću listu takvih, manje vidljivih, razlika:
| primitivni reptili | Sisari |
| nemaju fenestre u lobanji | veliki otvor (fenestra) |
| moždani okvir labavo vezan za lobanju | moždani okvir čvrsto vezan za lobanju |
| nema sekundarnog nepca | kompletno koščato sekundarno nepce |
| svi zubi isti | različiti sekutići, očnjaci i kutnjaci |
| obrazni zubi bez krune | obrazni zubi sa krunom |
| zubi se stalno zamenjuju novim | zubi se zamenjuju najviše jednom |
| svi zubi sa jednim korenom | kutnjaci sa dva korena |
| zglob desni povezuje quadrate i articular kosti | zglob desni povezan direktno sa kostima lobanje |
| donja vilica sačinjena od više kostiju | donja vilica sačinjena od jedne kosti |
| jedna kost u uvetu | tri kosti u uvetu |
| spojene spoljne nozdrve | odvojene spoljne nozdrve |
| jednostruka occipital condule(veza između lobanje i kičme) | dvostruka occipital condule |
| duga vratna rebra | kratka ili nepostojeća vratna rebra |
| postoje lumbarna rebra | nema lumbarnih rebara |
| nema dijafragmu | ima dijafragmu |
| udovi rašireni sa strane tela | udovi pravo ispod tela |
| ramena kost prosta | ramena kost sa masivnim izraštajem za vezu sa mišićima |
| kosti pelvisa razdvojene | kosti pelvisa spojene u jednu celinu |
| dve vertebre u pojasu kuka | tri ili više vertebri u pojasu kuka |
| raspored kostiju u prstima 2-3-4-5-4 | raspored kostiju u prstima 2-3-3-3-3 |
| telesna temperatura varira | telesna temperatura konstantna |
Zapamtite da su osobine navedene u levoj koloni osobine primitivnih reptila. Kasniji, razvijeniji reptili se ne uklapaju uvek u ovu sliku!
Priču počinjemo od primitivnih reptiloida sa značajnim brojem vodozemačkih karakteristika, kao što je bio u prethodnom poglavlju pomenuti Hylonomus (koji je živeo oko 315Ma). Ubrzo nakon njega se pojavljuju Proteoclepsydrops i Clepsydrops (cca. 310Ma), prvi sinapsidi, kod kojih se pojavljuje mala fenestra na lobanji.
Sledi Archaeothyris (cca. 300Ma), kod koga se lobanja malo modernizuje, mada je moždani okvir još uvek nepovezan sa ostatkom lobanje. Pojavljuju se prvi znaci različitih vrsta zuba. Telo je potpuno reptilsko, spolja čak liči na vodozemce (i još uvek ima vodozemačke osobine u građi vilice i nogu). Kuk počinje da se razvija i kost postaje veća i jača.
Varanops (cca. 290Ma) nije u liniji direktnih predaka, ali je veoma blizak srodnik: lobanja počinje da liči na sisarsku po mnogim karakteristikama. Moždani okvir počinje da se jače povezuje sa lobanjom, mišići vilice postaju snažniji i pomeraju se unapred, telo postaje uže i dublje. Kičma postaje snažnija. Razvoj kuka se nastavlja.
Rod Haptodus čini nekoliko srodnih vrsta reptila kod koje nastavljaju da se približavaju građi sisara. Veza između moždanog okvira i lobanje postaje još snažnija, vilica postaje snažnija, pojavljuju se značajne razlike u vrstama zuba. Dok sa jedne strane zglobovi i građa kičme postaju sličniji sisarima, sa druge strane na slici skeleta možete da vidite izražene reptilske karakteristike, kao što su vratna i lumbarna rebra. Umesto dosadašnje dve, tri vertebre postaju povezane u pojasu kuka.
Sphenacodon (270Ma) i srodne vrste idu još jedan korak dalje. Detalji lobanje postaju još sličniji sisarima. Razlike u zubima postaju izražene – jasno se razlikuju sekutići i očnjaci; ali zubi se i dalje stalno zamenjuju. Donja vilica je još uvek sačinjena od više kostiju, ali se pojavljuju prvi tragovi izrasline koja će kasnije biti ključni deo slušnog sistema kod sisara (za detalje o razvoju slušnog sistema pogledajte napomenu na kraju teksta).
Ovde dolazimo do terapsida, grupe vrsta na polovini puta između reptila i sisara. Biarmosuchia (260Ma) je jedan od prvih, najprimitivnijih terapsida. Još uvek su prisutne neke vodozemačke osobine (građa mišića vilice, na primer), mada se najveći atavizmi ipak gube. Gornja vilica Biarmosuchie, na primer, je spojena sa lobanjom, dok je kod vodozemaca, najprimitivnijih reptila, i svih iznad navedenih vrsta odvojena.
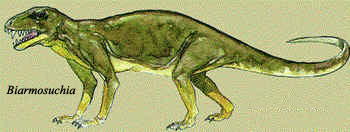
Fenestra raste dalje, i dostiže veličinu sličnu onoj kod sisara. Nastavlja se snažnije povezivanje moždanog okvira sa lobanjom. Gornja vilična kost (maksila) se razvija nagore, proizvodeći prve tragove primitivnih sisarskih sinusa. Mišići vilice počinju da se smanjuju, osim glavnog sisarskog viličnog mišića. Kosti prednjeg nepca se razvijaju unazad; mada ne formiraju potpuno sekundarno nepce, ovaj pravac razvoja je identičan sisarskom. Očnjaci postaju izraženi, zamena zuba varira; kod mnogih terapsida se pojavljuje ljudima dobro poznati raspored mlečnih zuba u mladosti, koji bivaju zamenjeni stalnim zubima kod odrasle životinje. Kičma se ne menja mnogo, ostajući uglavnom reptilska, sa rebrima celom dužinom tela. Prednji udovi postaju pokretljiviji, a u zadnjim udovima femur i pelvis postaju prilagođeniji za viši nivo odgurivanja od tla. Broj kostiju u prstima varira kod različitih terapsida: neki imaju reptilski, neki sisarski broj kostiju. Vertebre vrata i repa postaju vidljivo različite od ostalih. Pojavljuju se tragovi bubne opne u korenu vilice.
Procynosuchus (je prvi cinodont, grupa za koju neki paleontolozi smatraju da bi trebala da bude klasifikovana kao prvi sisari. Sekundarno koščato nepce je prisutno, ali još uvek nepotpuno. Reptilski mišići vilice postaju još manji, osim glavnog koji se razvija i postaje daleko veći; od ovog mišića će nastati ogromni sisarski vilični mišić. Smanjuje se broj sekutića u vilici – raniji reptili imaju šest, Procynosuchus ima četiri, prvi sisari imaju tri. Velike kosti vilice se smanjuju i nastavlja se napredak ka modernom slušnom sistemu; vidljivi tragovi bubne opne. Vilični zglob je još uvek potpuno reptilski. Kičma postaje sličnija sisarima: rebra u lumbarnom delu tela se smanjuju, a vrat postaje pokretljiviji. Mada su fosilni dokazi nedovoljno jasni da bi se moglo reći bez sumnje, neki paleontolozi smatraju da se kod Procynosuchusa pojavljuju i prvi znaci postojanja dijafragme. Kosti prednjih udova se produžuju i prilagođavaju za uspravniji hod. Konačno, kosti svih cinodonata poseduju takozvane Haversove kanale, koji se pojavljuju isključivo kod toplokrvnih životinja.
Trinaxodon (cca. 250Ma) je dobar primer sledećeg koraka u razvoju. Sekundarno nepce je skoro potpuno, diferencijacija zuba je završena mada svi zubi i dalje imaju samo jedan koren. Od tri kosti koje definišu donju vilicu, dve skoro nestaju, ostavljajući jednu dominantnom. Vilični zglob počinje da liči na sisarski. Veza lobanje sa kičmom je tačno na pola puta između reptilske i sisarske, sa dve odvojene površine, ali ne onoliko izdužene kao kod sisara. Lumbarna rebra su manja, kost ramena (scapula) pokazuje razvoj sisarskog ramenog mišića. Kosti udova su potpuno prilagođene za sisarski uspravni hod sa udovima ispod tela. Rep postaje kraći, građa rebara i kičme ukazuje da je skoro sigurno postojala dijafragma. Fosili odraslih i mladih životinja se nalaze zajedno, što je moguća indikacija da su Trinaxodoni aktivno pazili na svoju mladunčad.
Cynognathus (cca. 240Ma) je potpuno razvijeni cinodont koji je zanimljiv najviše zbog razvoja vilice i zuba. Kutnjaci imaju razvijenu krunu, i gornji zubi se uklapaju sa donjim. Donja vilica je 90% sačinjena od jedne kosti. Prisutna su dva vilična zgloba, reptilski i sisarski. Rebra su veoma blizu sisarskoj građi, udovi pravi su ispod tela. Po otiscima stopala se vidi da je Cynognathus imao krzno.

Diademodon (240Ma, period se preklapa sa Cynognathusom) nastavlja trend, sa još jačim mišićima vilice, razvijenijim krunama na zubima, i donjom vilicom koja je 98% jedna kost. Skoro potpuno razvijeno sekundarno nepce, ali i dalje prisutna dva vilična zgloba. Broj kostiju u prstima postaje sisarski 2-3-3-3-3, i ostaje takav od ove tačke nadalje. Lumbarna rebra postaju veoma mala, otvarajući više mesta za nedvosmisleno definisanu dijafragmu. Probelesodon (cca. 235Ma), sledeći u sekvenci, ima kutnjake sa dva korena, sekundarno nepce je još bliže konačnom izgledu kod sisara, lumbarna rebra potpuno nestaju.
 Diarthrognatus (209Ma) je predstavnik tritelodonta, razvijene grupe cinodonta. Sinusi postaju razvijeni, Eustahijeva tuba se pojavljuje između uveta i grla. Vilični zglob je još uvek dvostruk, ali sisarski zglob postaje potpuno razvijen. i igra glavnu ulogu u pokretanju; reptilski zglob zakržljava, i uglavnom igra ulogu u čulu sluha. Kičma vrata postaje još sličnija sisarskoj, što omogućava veću pokretljivost. Kuk i butna kost su skoro potpuno sisarski. Rame se razvija zajedno sa novim mišićem.
Diarthrognatus (209Ma) je predstavnik tritelodonta, razvijene grupe cinodonta. Sinusi postaju razvijeni, Eustahijeva tuba se pojavljuje između uveta i grla. Vilični zglob je još uvek dvostruk, ali sisarski zglob postaje potpuno razvijen. i igra glavnu ulogu u pokretanju; reptilski zglob zakržljava, i uglavnom igra ulogu u čulu sluha. Kičma vrata postaje još sličnija sisarskoj, što omogućava veću pokretljivost. Kuk i butna kost su skoro potpuno sisarski. Rame se razvija zajedno sa novim mišićem.
Sinoconodon (208Ma) je prvi potpuno prihvaćeni proto-sisar. Lice je potpuno sisarsko, mozak je još razvijeniji. Vilični zglob je skoro potpuno sisarski – reptilski zglob je još uvek prisutan, ali je sitan i zakržljao.
Sledi čitava grupa vrsta poznata kao morganukodoni, jedna od kojih je bio Haldanodon (205Ma). Sekundarno nepce je skoro potpuno, zubi su potpuno sisarski osim par detalja u građi kutnjaka. Sistem zamene zuba je takođe potpuno sisarski. Dno moždanog okvira postaje povezano sa lobanjom. Ušni aparat, sastavljen od minijaturnih ostataka reptilskog viličnog zgloba, je praktično potpuno razvijen. Svi ostali aspekti skeleta su sisarski.
Biolozi smatraju morganukodone prvim pravim sisarima. Ova grupa je živela prilično rašireno u periodu Jure, kada su dinosaurusi bili na svom vrhuncu.
Poslednji reptilski detalji pomenuti kod morganukodona nestaju kod pantotera i drugih trikonodonata, grupe koja preuzima primat među sisarima tokom zadnjeg dela Jure i tokom perioda izumiranja dinosaurusa. Nepce postaje potpuno, struktura kutnjaka postaje jasno definisana, ušni aparat je kompletiran…i sa time, i naš pregled evolucije sisara od reptila, korak po korak.
Kratak pregled razvoja sisara
Nakon izumiranja dinosaurusa, sisari popunjavaju napuštene ekološke niše jednu po jednu, i broj vrsta raste eksponencijalno. Postoje na desetine hiljada prelaznih fosila, i većina grana razvoja (osim šišmiša) je veoma dobro dokumentovana. U okviru ovog teksta, mi ćemo obraditi samo granu – onu koja vodi ka primatima, direktnim precima čoveka. Ali pre nego što se zaputimo na taj put, vredi pogledati makar samo najopštije tačke ukupne evolucije sisara.
Najraniji sisari koje smo pomenuli iznad su imali krzno, skoro sigurno su imali primitivne mlečne žlezde, ali su i dalje legli jaja umesto žive mladunčadi. Ovi primitivni sisari su se još pre izumiranja dinosaurusa razdvojili na četiri reda.
Prvi red, monotreme, je zadržalo većinu ovih primitivnih osobina. Najraniji siguran pripadnik monotrema, Steropodon galmani je živeo pre oko 140Ma, i pokazuje seriju karakteristika koje su nasledili svi njihovi kasniji potomci. Do danas su preživele samo dve vrste monotrema, kljunar i ehidna, i one i dan danas legu jaja, doje mladunce koji se iz tih jaja izlegu mlekom iz veoma primitivnih mlečnih žlezda, i poseduju mozak koji je znatno manje razvijen u poređenju sa višim sisarima.
Druga grupa su multituberkulati, sa zubima koji pokazuju karakteristične redove malih vrhova. Multituberkulati su jedina glavna grana sisara koja je danas potpuno izumrla: postojali su od polovine perioda Jure (170Ma) do početka Oligocena (cca. 34Ma). Poznato je oko osamdeset rodova ovih životinja, koje su po veličini i izgledu bile donekle slične današnjim glodarima
Treći i četvrti red su povezani osobinom koja definiše više sisare: rađanje živih mladunaca. Rađanje živog mladunčeta nije neki poseban podvig sam po sebi – dovoljno je da životinja zadrži jaje u sebi do trenutka kada ono treba da se izlegne. Evolutivna prednost je ogromna, pošto majka može da nastavi da traži hranu ili lovi umesto da sedi na jajetu i čeka da se ono izlegne. Problem koji se javlja je nešto sasvim drugačije: ako mladunče raste dovoljno dugo u telu majke, majčin imuni sistem će ga napasti i uništiti.
Treći red, Marsupialia Marsupialia ili torbari, se sastoji od životinja kod kojih se prvi deo razvoja mladunčeta odvija unutar organizma, ali mladunče biva rođeno pre nego što bi majčin imuni sistem imao vremena da reaguje. Prvobitni torbari su najverovatnije ostajali pored novorođenih mladunaca da ih doje (što je i dalje mnogo bolje nego da se prvo sedi na jajetu, pa onda povrh toga ostaje pored mladunčeta), ali se ubrzo javlja jedna tipično torbarska osobina: mladunci se rađaju sa razvijenijim prednjim udovima, pomoću kojih se drže za majčino krzno dok sisaju, što omogućava majci da nastavi potragu za hranom i u toku dojenja. Kod nekih kasnijih, razvijenijih torbara se javlja prevoj u koži sličan “torbi” (po čemu su i nazvani) u kome mladunče može udobno da leži, ili čak i spava, dok se majka kreće.
Evolucija placente
Male su šanse da se može naći fosil placente, koja je meko tkivo koje je prisutno samo kod trudnoće. Otud mi nemamo fosilne dokaze da se placenta razvila od membrane jajeta primitivnijih reptila; ali imamo strukturalne i genetske dokaze koji to pokazuju. Konkretno, geni koji učestvuju u signalnim putevima koji definišu placentu su kopija genetskih sistema koji inače upravljaju razvojem membrane jajeta, a osnovni strukturalni proteini su identični. Takođe, razvoj placente od relativno jednostavne membrane do kompleksne strukture kakvu vidimo kod viših sisara se može pratiti kroz poređenje placente različitih vrsta sisara danas.
U razumevanje evolucije placente se ulažu velike količine novca i truda, pošto mnogi placentalni poremećaji (koji vode do pobačaja i drugih problema u trudnoći) imaju korene u nestabilnim sistemima koje je evolutivni proces proizveo.
Fosilni zapis torbara je veoma bogat, počevši od prvih tragova torbarske građe kod Pariadens kirklandi (cca. 95Ma), preko prvih tragova torbe kod Sinodelphys szalayi (cca. 80Ma), do potpunih torbarskih osobina gigantskog Diprotodon optatuma. Mada su torbari danas preživeli samo u Americi i Australiji, fosilni ostaci njihovih predaka su nađeni i u Africi, Aziji pa čak i na Antarktiku.
Konačno, četvrti red je Eutheria, placentalni sisari. Kod njih se unutrašnja membrana jajeta razvija u placentu, membranu koja razdvaja embrion od majčinog imunog sistema. Pošto je placenta meko tkivo, njen razvoj ne može da se prati pomoću fosila direktno, već samo indirektno pomoću muskuloskeletalnih osobina koje su specifične za placentalne sisare.
Vincelestes neuquenianus (cca. 135Ma) je najraniji poznati fosil čija lobanja pokazuje osobine koje imaju samo placentalni sisari i torbari; ali, na žalost, telo nije očuvano, tako da su zaključci nesigurni. Skoro je sigurno da je početak razdvajanja ova dva reda počeo u ovom periodu, pošto nekih desetak miliona godina kasnije nalazimo niz vrsta koji pokazuju razdvajanje ova dva reda.

Prvi nedvosmisleni predak torbara je Sinodelphys szalayi (125Ma), kod koga vidimo dalje prilagođavanje karlice, i promene na mnogim kostima udova koje su tipične za torbare. Sledi Deltatheridium (80Ma, vrhunac doba dinosaurusa), koji pokazuje ne samo dalji razvoj torbarskih osobina, vec i razvoj donjih udova ciji ekstrem vidimo kod današnjih kengura. Nakon Deltatheridiuma sledi oko dvadeset poznatih prelaznih fosila (Sulestes, Kokopellia, Pediomys, Anchistodelphys…) do današnjih formi torbara.
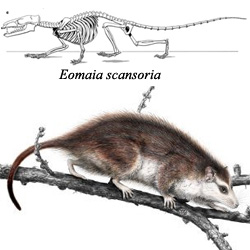
Prvi nedvosmisleni predak placentalnih sisara je Eomaia scansoria (125Ma). Kosti karlice počinju da se adaptiraju za rađanje živih mladunaca, ali je prolaz još uvek suviše mali; Eomaia je verovatno rađala veoma malu i nerazvijenu mladunčad, na koje je zatim morala da pazi (što se uklapa u predviđenu shemu razvoja). Eomaju sledi niz sisara sa sve vidljivijim prilatođavanjima za rađanje živih mladunaca, kao što su Kennelastes i Asioryctes, pa zatim insektivore koje su živele u periodu izumiranja dinosaurusa, kao što su Cimolestes ili Procerberus.
Od sisara do hominida
Fosilni zapis ranih primata je relativno slab, mada noviji nalazi iz Španije i severne Afrike polako počinju da popunjavaju nedostajuće informacije. Podela primata je, međutim, promena na mnogo manjem nivou od onih o kojima smo diskutovali iznad (broj i veličina promena između ranih sisara i primata je daleko manja nego između ranih reptila i sisara), a u rekonstrukciji nam pomaže činjenica da su mnoge primitivne vrste primata opstale do danas, tako da možemo da vidimo prelazne korake uživo
Gorepomenute insektivore su se rano podelile na dve grane. Sa jedne strane su se odvojili preci današnjih glodara, sa druge strane male, mišolike životinje koje su živele u drveću (Ptilocercus je jedan primer). Uporedimo osnovne osobine ovih insektivora sa osobinama primata:
| insektivore | primati |
| dugačak, ali slab rep | dugačak, mišićav rep |
| kandže | nokti |
| zubi i vilica slične glodarima | primatski zubi i vilica |
| oči sa strane glave | oči napred u licu |
| dugačka njuška | kratka njuška |
| očna duplja otvorena | postorbitalni most zatvara očnu duplju |
| osrednji mozak sa razvijenim olfaktornim centrima | veliki mozak, sa manje razvijenim olfaktornim centrima, ali veoma razvijenim vizuelnim centrima |
| očnjaci izbačeni sa strane | vertikalni očnjaci |
Smilodectes pokazuje uticaj života u granama drveća. Stopala i šake postaju razvijenija, mišićavija i sa dužim prstima, kako bi životinja lakše mogla da se drži za granu. Rep postaje mišićaviji i veći, pošto služi za kontrolu pri skakanju sa grane na granu (sličan razvoj repa vidimo kod današnjih veverica). Drveće je trodimenzionalna životna sredina: procena razdaljine je od ključne važnosti. Oči Smilodectesa se pomeraju blago unapred, mada su još uvek sa strane glave. Njuška se smanjuje. Olfaktorni centri (koji kontrolišu njuh) se smanjuju, dok se istovremeno povećava vizuelni korteks (koji upravlja čulom vida).
Adapis predstavlja sledeći korak (poznato je nekoliko vrsta u ovom rodu, sa različitim nivoima adaptacije). Rep postaje još snažniji, čulo vida se dalje razvija, oči se pomeraju još više unapred. Pojavljuje se postorbitalni most, očnjaci postaju uspravniji. Pelycodus je prvi poznati primat sličan modernim lemurima. Kandže se skraćuju i postaju sličnije noktima. Zubi i vilica su još uvek slični glodarima i insektivorama (ovo se može izraženo videti u Aye-Aye, vrsti primitivnih primata koja je preživela do današnjih dana na Madagaskaru).

Ovde dolazi do podele. Neke vrste slične Adapisu zadržavaju njegove primitivne osobine, i njihovi potomci, lorisi i lemuri, i dan-danas imaju njegovu vilicu sa glodarskim osobinama. Sa druge strane, odvaja se grupa poznata kao Haplorhini. Oči postaju veće, i potpuno okrenute unapred. Mozak postaje još veći u odnosu na veličinu tela, lice postaje manje, njuška se povlači.
Primer vrste iz ovog perioda je Necrolemur, ali postoje i živi predstavnici: tarsijeri, u javnosti malo poznata familija primata koja danas opstaje samo na nekim ostrvima u Pacifiku.

Razdvajanje kontinenata dovodi do razdvajanja primata. Primati koji ostaju izolovani na Američkom kontinentu su danas poznati kao Plathyrrhini, i postaće preci današnjih američkih majmuna. U Evropi, Aziji, i Africi ostaje grupa poznata kao Catharrini.
Plathyrrhini će u izolaciji razviti mišiće repa do tačke na kojoj mogu da hvataju predmete njime, njihove ruke će se razviti za kretanje kroz ljuljanje sa grane na granu, i živeće isključivo u tropskim zonama. Ako prosečnog čoveka danas pitate da opiše majmuna, šanse su da će opis odgovarati Platirinima. Platirini nisu preci čoveka.
Catharrini su značajno drugačiji od svojih američkih rođaka. Njihov rep ostaje relativno nerazvijen – ne mogu njime da hvataju predmete ili da pomocu njega vise sa drveta. Mozak i socijalno ponašanje su, međutim, daleko razvijeniji, sa kompleksnim hijerarhijama, vezama, i sistemima društvenih obaveza.
Negde u toku ranog Oligocena se događa sledeći ključni razlaz: Catharrhini se dele na pretke današnjih Afričkih i Azijskih majmuna sa jedne strane, i hominide sa druge strane. Detalji ove podele su na žalost veoma magloviti: u ovom periodu dolazi do nagle promene klime (globalno zahlađenje), koje dovodi do masovnog izumiranja mnogih vrsta, i naglog smanjenja broja primata (što automatski smanjuje i šanse njihove fosilizacije).
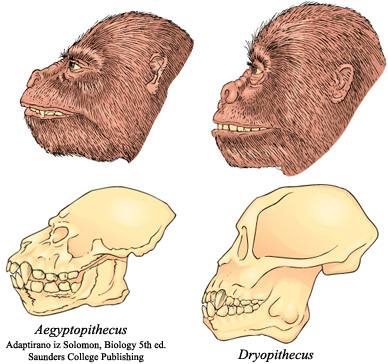
Aegyptopithecus je jedan od najranijih katarina, i pokazuje sve njihove osobine. Vilica gubi zadnje glodarske osobine, i postaje sličnija vilici hominida (mada je još uvek više izbačena unapred). Rep počinje da se smanjuje kako životinja postaje sve veća i sve manje zavisna od repa pri kretanju.
Proconsul africanus je prvi direktan predak hominida. Rep potpuno nestaje, kutnjaci poprimaju standardni oblik sa pet vrhova i procepom u obliku slova Y. Pokretljivost ramena se povećava. Zglobovi na rukama i građa kičme još uvek slični majmunima.
Kod Dryopithecusa se ove osobine ustaljuju, i telo poprima standardni hominidski izgled. Zglobovi se adaptiraju za hominidski “dve noge potpomognute rukama” način kretanja. Na slici se može jasno videti dramatičan razvoj lobanje od veoma majmunolike do proto-hominidne, kao i završetak uklanjanja preostalih osobina koje karakterišu primitivne primate.
Zadnja podela u okviru ovog teksta je podela hominida neposredno posle Dryopithecusa: sa jedne strane se odvaja linija Sivapithecusa i Ramapithecusa, koja prelazi u Aziju i od koje nastaju današnji orangutani. Sa druge strane se odvajaju Kenyapithecusi. Primeri vrsta iz ovog prelaza su Sahelanthropus tchadensis i Ardipithecus ramidus – čija je priča, kao i dalja evolucija ljudi, obrađena u tekstu o evoluciji čoveka.
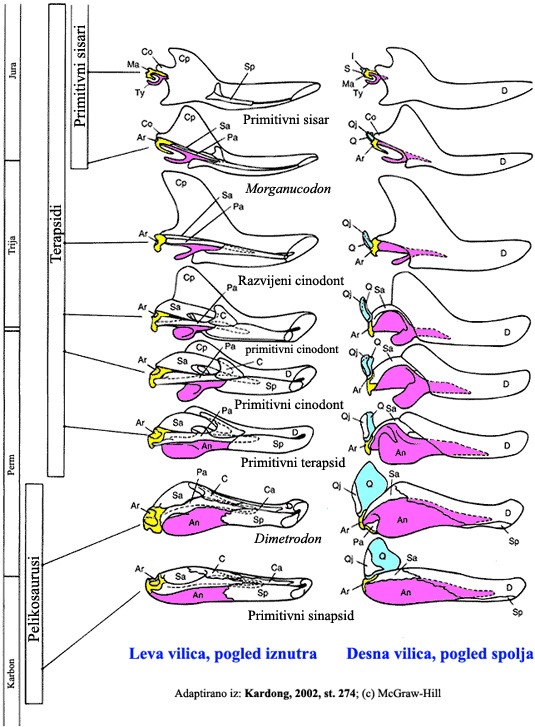
Napomena 1: Slušni aparat i reptilska vilica
Razvoj slušnog aparata sisara je fantastičan primer evolucije, gde se može videti postepeni prelaz, korak po korak, do konačnog oblika. Pritisak koji je proizveo ovu evoluciju je zasnovan na kvalitetu sluha. Naime, prvobitni primitivni vodozemci su bili potpuno gluvi kada su izašli na tlo: mogli su da osete samo vibracije koje su se kretale kroz tlo, i vrlo niskofrekventne (duboke) tonove. Sa svakim korakom smanjenja slušnih kostiju, organizam bi postajao sve osetljiviji na više frekvencije; dobar sluh, uporediv sa onim kakav danas imaju ljudi, se razvio tek kod morganukodona.
Pogledajmo sekvencu fosila koji ovo pokazuju (imate u vidu da između svih prikazanih koraka postoje međukoraci; zbog prostora na slici, ovde su prikazani samo glavni prelazi).
Napomena 2: Krzno
Dlake se retko i loše fosilizuju, tako da nam fosilni zapis daje manje informacija kako se ono razvilo (mada nam otisci pokazuju da su već cinodonti posedovali krzno). Dodatne odgovore na ovo pitanje možemo da potražimo u genetskim i fiziološkim dokazima.
Razvoj krzna nije neka posebna genetska misterija: dlake su sačinjene od keratina, osnovnog strukturalnog proteina kože koji je počeo da se izdužuje van njene površine.Ali koji je evolutivni pritisak učestovao u ovome? Krljušti koje su reptili nasledili od vodozemaca su sasvim dobar oblik kože što se tiče zaštite od spoljne sredine – možda čak i previše dobar. Naime, čulo dodira slabo funkcioniše kroz debeli sloj krljušti, što ograničava sposobnost organizma da se orijentiše pomoću dodira.
Prve dlake su verovatno rasle između krljušti, i služile kao organi dodira: pošto sinapsid nije mogao da prepozna dodir pomoću kože, prepoznavao ga je pomoću dlaka. Ovaj sistem je opstao sve do danas: i ljudi mogu da osete dodir pomoću dlaka na telu, a mnogi sisari imaju posebno razvijene organe zasnovane oko osetljivih dlaka (mačji brkovi su dobar primer). Neki sisari koji i dalje imaju strukture na koži slične krljuštima i dan danas koriste dlake koje rastu između krljušti za čulo dodira (armadilo, na primer).
Kada su se jednom razvile, dlake su se verovatno (bar po kalkulacijama molekularnih biologa) brzo razvile u mehanizam za kontrolu temperature: naši hladnokrvni ili nepotpuno toplokrvni preci su tako mogli da održe toplotu tela na aktivnom nivou mnogo duže nego inače.
[prethodno poglavlje][nazad na vrh strane][sledeće poglavlje]