
- Šta je uopšte evolucija
- Evolucija danas: mutacije i nastanak informacija
- Evolucija danas: selekcija i širenje informacija kroz populaciju
- Evolucija danas: prirodna selekcija, geni i populacije
- Teorija evolucije života na Zemlji
- Dokazi: Evoluciona istorija i ugnježdena hijerarhija
- Dokazi: Genetika, genomika i tragovi evolucije
- Dokazi: Fosilni zapis i razvoj života
- Dokazi: Razvojni i fiziološki tragovi
- Zakoni prirode ili slučajna konfluenca dokaza?
Organizmi se razvijaju na osnovu genetskog koda u okviru njihove DNK. Otud, lako je napraviti grešku, i zamisliti DNK kao neku vrstu “plana” ili “mape” organizma – u kojoj su geni onda neka vrsta instrukcija “kako sagraditi živo biće”.
U stvari, DNK ne sadrži takve informacije uopšte. Nigde u DNK nije napisan, recimo, oblik srca nekog organizma; ne postoji čak ni informacija da ovaj organizam uopšte ima srce. U ljudskoj DNK nigde nema opisa kičme, mozga, želuca, jetre, kože…
Umesto toga, geni funkcionišu drugačije: većina gena sadrži u sebi informacije o pojedinim proteinima, kao i o tome kada, gde i koliko ti proteini treba da se proizvode. Na osnovu ovih proteina, početna odvojena jajna ćelija se deli u dve, te dve se dele u četiri, četiri u osam ćelija…ali ne jednako. U nekim od ovih ćelija, ovi proteini će ugasiti određene gene, u drugima će uključiti neke druge gene. Ovo onda menja način na koji se ćelije dele, menja njihov sadržaj, i menja njihovu funkciju.
Rezultat ovoga je razvoj organizma; a nuzprodukt je njegova modularnost. Naime, radi pojave neke nove osobine – nove kosti, novog uda, ili novog organa – nije potreban istovremeni razvoj nerava, krvnih sudova, i svih potrebnih tkiva. Sve to se proizvodi automatski zajedno sa novorazvijenim organom. Pogledajmo ovo na jednom malom primeru.
U toku razvoja ptičjeg embriona, dva mala nabora se pojavljuju sa strane. U okviru ovih nabora, između drugih stvari, je parče tkiva zvano ZPA, u kome su određeni proteini aktivni. Ovo tkivo počne da se razvija u dinosaursku prednju šapu: grupu standardnih kostiju tetrapodskog prednjeg uda, uključujući tu i početak kostiju koji formiraju prste. Nedugo zatim, međutim, dodatni signali menjaju pravac razvoja, i umesto šake ili šape, dobijamo krilo (slika ispod, pod A; ostale kosti prednjeg uda – humerus, fibija i tibula – ostaju iste).
Ilustrativan je jedan eksperiment koji se ovde može izvesti. Ako mikrohirurški isečemo nekoliko ćelija iz ZPA zone i presadimo ih na drugo mesto, one nastavljaju da se razvijaju – i proizvode dodatnu kopiju cele “šake”, tj. celog vrha krila (slika ispod, pod B).
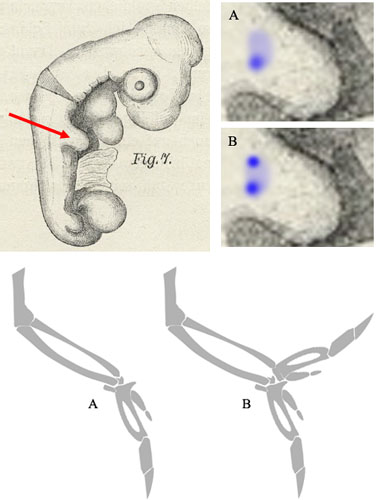
Crtež pilećeg embriona iz knjige Viljema Hisa, jednog od ranih embriologa. Skica kostiju krila adaptirana od L. Shyamal.
Ovo dodatno krilo će automatski biti prokrvljeno krvnim sudovima, imaće nerve koji prenose osećaje dodira, toplote, hladnoće, bola…imaće čak i mišiće, koje ptica može svesno da kontroliše.
Ovo važi za sve delove tela. Uzmimo oko: za funkciju oka je potrebno više stotina gena. Ali razvoj i pozicija oka zavisi od jednog kontrolnog gena; ako njega uklonimo, rađa se životinja bez očiju. No, ako promenimo tkivo u kome je ovaj gen aktivan, možemo time promeniti mesto na kome će izrasti oči. U eksperimentima su tako proizvedene vinske mušice kojima dodatne oči rastu na kolenima, ili na antenama, prostom promenom jednog gena.
Mada ovakva stvorenja ne bi dugo preživela u prirodi, i nisu primer evolucije, ona jesu primer kako se oblik organizma može menjati kroz male modifikacije razvojnih gena. Proizvodnja krila od prednje šape dinosaurusa nije zahtevala ogromne izmene u genetskom kodu: naprotiv, male promene u regulatornim genima proizvode postepene promene u obliku i rastu kostiju, mišića i nerava – koordinisano.
Isto važi i za veliku većinu svih ostalih promena u toku evolucije životinja. Promene u osnovnoj biohemiji su veoma, veoma retke: najveći deo promena potrebnih da riba postane vodozemac, da vodozemac postane sisar, ili da sisar postane čovek je bazirana na promenama gena koji regulišu razvoj.
Važno je razumeti da su ovi geni evoluciono veoma stabilni. U razvoju vrste, gen koji određuje razvoj jedne strukture može postati osnova za dalju nadogradnju. Recimo, gen SEMA4A proizvodi signal na osnovu koga se usmerava rast određenih nervnih ćelija; ali ovaj isti signal su (evoluciono kasnije) počele da koriste i druge ćelije, u okviru razvoja određenih kostiju i imunog sistema. Ako SEM4A mutira tako da proizvede pozitivnu promenu u pravcu rasta nervnih ćelija, šansa je da će pritom efekti na razvoj kostiju i imunog sistema biti negativni. Otud, praktično bilo kakva mutacija biva eliminisana iz populacije, i pozitivni razvoji na jednom planu su onemogućeni negativnim razvojima na drugim.
Ovo nije apsolutna prepreka za evoluciju, koja može ovaj problem da zaobiđe. Ali ovo znači da se sama struktura uvreženih razvojnih gena veoma teško menja: pošto praktično sve promene dovode do eliminacije jedinke koja nosi promenjeni gen, jedina bića koja ostavljaju potomstvo su ona koja nose nepromenjenu verziju. Najosnovniji razvojni geni u okviru životinja su često ostali toliko nepromenjeni da su danas praktično jednaki kod sisara i insekata.
Lep primer za ovo je upravo gen za razvoj oka: naučni su uništili ovaj gen kod mušica, i dobili su slepe mušice. Ali onda su uzeli gen za razvoj oka iz miševa, ubacili ga u genom tih mušica – i oko mušice se razvilo potpuno normalno.
Dakle, isti gen koji kod miša dovodi do razvoja standardnog kičmenjačkog oka (koje je praktično isto kao i ljudsko), kod mušice može da pokrene nastanak mušičjeg kompozitnog oka (koje je sastavljeno od stotina odvojenih šestougaonih sočiva).
Genima kao što su ovi se bavi grana nauke poznata kao evoluciona razvojna biologija (engleski “evolutionary developmental biology”, zbog čega je šire poznata po nadimku “Evo Devo”). No, zašto je reč “evoluciona” uključena u ovo ime?
Prvi razlog je nivo kompleksnosti razvojnih puteva u organizmu. Ako se jedan organizam posmatra individualno, veoma je teško razlučiti šta se i gde tačno događa u pojedinačnim ćelijama u toku razvoja. Ali ako se posmatra evolutivno nasleđe, možemo pojavu osobine da pratimo na genetskom nivou, i da ustanovimo koji geni su za nju odgovorni – i tako, možemo da prepoznamo koji geni kod čoveka vrše istu funkciju (i možda igraju ulogu u nekoj bolesti).
Na primer, određen broj ljudske dece se rađa svake godine sa anomalijom u građi srca: komora srca nije potpuno podeljena, već omogućava mešanje arterijske i venske krvi, što proizvodi razne probleme. Da bi ustanovili koji geni su odgovorni za ovaj razvoj, naučnici su se okrenuli vodozemcima (koji imau jednodelnu srčanu komoru), kornjačama (koji imaju delimično podeljenu komoru) i krokodilima (koji imaju potpunu podelu). Zajednički preci ove tri vrste su bili veoma srodni ranim sinapsidima, koji su preci svih sisara (pa samim tim i čoveka).
Koristeći evolutivne profile i analizirajući uporedan razvoj i genetsku aktivnost ćelija srca ovih životinja, naučnici su ubrzo identifikovali kontrolni gen po imenu Tbx5. Negativne mutacije u ovom genu, ili spoljni uticaji koji menjaju njegovu aktivnost, dovode do problema sa razvojem srca kod dece. Sada, na osnovu ove evolucione analize, može se početi sa razvojem ciljanih terapija usmerenih za sprečavanje i lečenje ovog čestog srčanog defekta.
Drugi razlog je evoluciona istorija ovih procesa. Da bi smo razumeli proces, moramo pre svega da razumemo početnu tačku i završnu tačku. Otud, recimo, da bi smo razumeli proces razvoja krila, moramo da razumemo njegovo evoluciono poreklo od šape dinosaurusa. U suprotnom, procesi koje posmatramo izgledaju potpuno arbitrarno i besmisleno.
Pogledajmo nekoliko konkretnih primera u kojima nasleđena osnova diktira dalje razvojne procese. Počnimo sa jednim čuvenim primerom, vratnim vertrebrama žirafida.
Primer: Vrat i nervi žirafe. Kada je ovaj tekst prvi put pisan, prelazni fosili žirafida (grupe u koju spadaju moderne žirafe) su još uvek bili retki i nejasni. Nova otkrića, uključujući i fosile izumrle vrste Decennatherium rex koji su nađeni tek 2017-te godine, su nam u međuvremenu dali potpuniju sliku (studiju ovog novog fosila možete pročitati ovde).
Svi sisari, od miševa do kitova, imaju sedam pršljenova u vratu. U fosilima predaka žirafe takođe vidimo sedam pršljenova. Ovo je lep primer fiksirane osobine u razvoju – pošto je razvoj kičme usko povezan sa razvojem mnogih drugih osobina, on se evolutivno jako teško može promeniti. Otud, ako se vrat žirafe razvio kroz nasleđe sa modifikacijom, možemo da očekujemo da ova osobina ostane nepromenjena. Pogledajmo sada kostur žirafe:
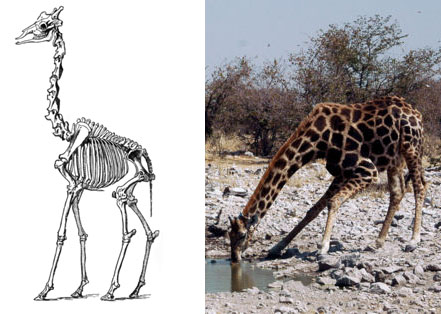
Bez obzira na činjenicu da im je vrat ogroman, današnje žirafe i dalje imaju samo sedam pršljenova u vratu. Ovi pršljenovi pokazuju delimične adaptacije – recimo, veza između njih je prerasla u pun zglob koji omogućuje da se vrat fleksibilnije kreće u stranu. No, ograničen broj pršljenova proizvodi čitav spektar problema, kao što vidimo na fotografiji žirafe koja pije: nezgrapna i ranjiva pozicija u kojoj se životinje nalaze kada god piju vodu ili pasu travu.
Drugim rečima, ne vidimo nikakve promene namenjene izduženju vrata. Ne vidimo planirano građenu strukturu. Umesto toga, vidimo standardnu ugnježdenu hijerarhiju, promenu usađenih nasleđenih osobina. Bez obzira koja je funkcija potrebna, osobina ne može da se prosto pojavi ili napravi, već može samo da se adaptira iz prethodno postojećih osnova.
Ovo je možda još lepše ilustrovano kroz jedan još stariji zaostatak evolucije: rekurentni laringijalni nerv. Pogledajmo malo evolucione istorije.
Dužina vrata i proporcije žirafa
Pored svih napomena, vredi dodati da žirafin vrat u stvari nije neverovatno dugačak u poređenju sa veličinom njenog tela; nama vrat žirafe izgleda impresivno uglavnom zato što se radi o gigantskoj životinji. Mnoge druge vrste, na primer gerunuk, imaju proporcionalno dugačak vrat, koji nama izgleda kraće samo zato što je cela životinja mnogo manja.
Kod riba, razvoj prednjeg dela tela u velikom delu zavisi od signala koje šalju škržni lukovi. Kada se embrion ribe razvija, tkivo škrga se pojavljuje vrlo rano, i svaki njegov sloj šalje određene signalne molekule, na osnovu kojih se onda orjentišu ćelije drugih tkiva. Tako, na primer, peti škržni luk oslobađa signal pomoću koga se ćelije krvnih sudova navode i formiraju šesti arterijski luk (krvni sud koji prati škrge). Nerv koji kontroliše mišiće i osećaj petog škržnog luka raste iz korena mozga pravo ka svojoj meti, prateći isti signal:
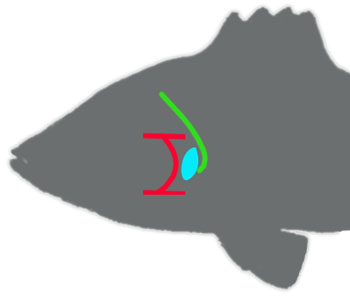
Deo anatomije riba: peti škržni luk je prikazan kao plavi oval; šesti arterijski luk je prikazan crvenim; četvrta grana vagusa (kod kasnijih tetrapoda rekurentni laringijalni nerv) je prikazana zelenom bojom.
U toku dalje evolucije, ovo tkivo je zadržalo svoju važnost. Embrioni svih tetrapoda (reptila, ptica, sisara) prolaze kroz takozvani “faringula” stadijum u razvoju, kada se blizu glave embriona formiraju lučne strukture na osnovu kojih se orjentiše razvoj ostatka prednjeg dela tela.
Deo hrskavičavog tkiva koji kod riba postaje peti škržni luk, u telu sisara postaje larinks, deo grla koji je važan za pravilno disanje i pravilnu proizvodnju zvukova. Krvni sudovi koji kod riba postaju šesti arterijski luk, pritom, kod sisara formiraju važan srčani krvni sud, poznat kao “ductus arteriosus”.
No, ovde imamo jedan mali, malecni problem. U toku evolucije sisara, srce je ostalo u okviru grudnog koša, dok su se glava i vrat izdužili. Međutim, onaj osnovni signalni sistem je ostao nepromenjen. On bi mogao da se promeni planski, ali evoluciono je njegova promena blokirana!
Vaša unutrašnja riba
Uopšte, razvojni sistemi cele lobanje su dobrim delom nasleđeni od ribljih. Razvoj ljudskog grla, nosa i vilice kontrolišu isti sistemi koji proizvode škržne kosti kod riba. Ljudski embrion u toku svog razvoja prvo prati ovaj riblji program, proizvodi osnovna tkiva za proizvodnju škržnih kostiju (sa svim nervima i krvnim sudovima koji su za to potrebni), pa tek zatim te strukture menja u današnje prepoznatljive ljudske oblike. Za više detalja, pogledajte knjigu Nila Šubina, “Riba u vama.”
Zašto? Razmislimo ovako: ako se desi mutacija zbog koje nerv izgubi sposobnost praćenja signala, on onda nikada neće stići do grla, i grlo neće funkcionisati kako treba. A ako se desi mutacija u signalnom sistemu koji navodi nerv, tako da se nerv sada kreće na drugačiji način, ta mutacija će istovremeno dovesti do deformacije srca i smrti od kardiovaskularnih problema. Otud, iako se struktura grla i tela drastično promenila, ovaj nerv nastavlja da prati isti signal i isti put ka svom cilju.
Ovo dovodi do situacije koju nalazimo kod svih tetrapoda, uključujući tu i čoveka. Rekurentni laringijalni nerv prati signal na osnovu koga silazi od korena mozga niz vrat sve do grudnog koša, zatim zaokreće uz srce oko aorte (i ductus arteriosusa, preostatka šestog arterijalnog luka), i onda se penje nazad uz vrat do larinksa.
Zbog praćenja signalnog puta koji je ostao kroz evoluciju još od riba, nepotrebna dužina ovog nerva je veliki problem za naše današnje hirurge (slučajno oštećenje ovog nerva u toku operacija na srcu, recimo, može da dovede do problema sa grlom i gubitka sposobnosti govora).
U vratu žirafe, ovo preuzima gigantske razmere: da bi povezao dve tačke udaljene samo nekoliko centimetara, rekurentni laringijalni nerv putuje više od šest metara.
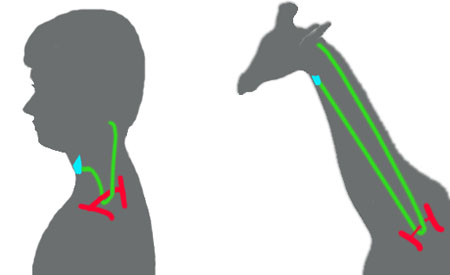
Ovo je samo jedan trivijalan primer ovakvog genetskog ostatka; slični primeri nasleđenih osobina koje danas samo proizvode probleme se mogu naći kod svih živih bića.
No, može se reći da su ovo samo male i direktne stvari, ograničene na jednu sporednu osobinu. Pogledajmo jedan primer mnogo dubljeg i mnogo starijeg problema.
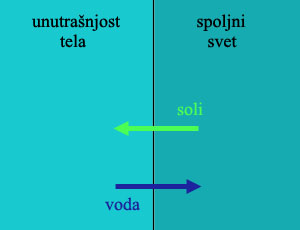
Primer: Evolucija bubrega. Osmotski pritisak je jedna od osnovnih činjenica fizike i biologije. Ako polupropustljivu membranu (kao što je ćelijska membrana životinja) stavimo između dve tečnosti sa različitim koncentracijama soli, dobićemo kretanje čestica. Čestice soli će se kretati iz slanije vode prema manje slanoj, dok će se voda kretati od manje slane ka više slanoj strani. Ovaj tok će se nastaviti dogod postoji ikakva razlika.
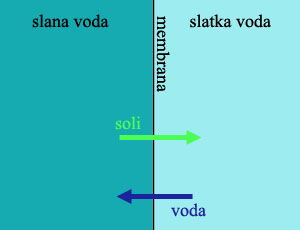
Koncentracija soli u ćelijskoj tečnosti je veoma važna za život. Da bi se procesi života mogli odigravati kako treba, potrebno je prisustvo natrijuma, kalijuma, hlorida, magnezijuma, kalcijuma, i mnogih drugih jona.
Za živa bića u moru, ovo nije problem. Morska voda sadrži sasvim dovoljnu količinu soli za održanje života, i sve što organizam treba da uradi je da dopusti ovim solima da se kreću iz okolne vode u njegovo telo. Praktično svi morski beskičmenjaci na svetu žive na ovaj način: njihove telesne i ćelijske tečnosti sadrže istu količinu soli kao i okolna morska voda.
Životinje u slatkoj vodi, međutim, imaju ozbiljan problem. Da bi mogle da uzimaju kiseonik iz vode, one moraju da drže svoje propustljive škrge u stalnom kontaktu sa okolnom tečnošću, u kojoj ima mnogo manje soli nego u telu. Otud, po pravilima osmoze, kako kiseonik ulazi u telo ribe kroz škrge, istovremeno u telo ulazi i okolna voda dok dragocene soli izlaze iz tela. Rezultat ovoga je da slatkovodne životinje nikada ne piju vodu, uvek traže izvore soli i minerala u svojoj ishrani, i dobar deo energije troše na neprestano pumpanje minerala i soli nazad u ćelije.
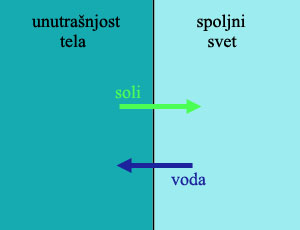
No, pored ovoga, one moraju da izbace iz tela suvišnu apsorbovanu vodu. One ovo čine u vidu urina, u kome istovremeno izbacuju i rastvorljive otpadne materije. Za ovu funkciju, evoluirali su posebni organi, bubrezi, koji troše značajnu količinu energije da iz tela (suprotno osmotskom pritisku) ispumpaju što više vode mogu, proizvodeći veoma redak urin.
Mada je ovo veoma značajan selektivni problem za slatkovodne organizme, on je takođe univerzalan: sve slatkovodne životinje (crvi, anelidi, insekti, ribe, vodozemci…) imaju isti problem, i moraju da ulažu istu količinu energije u ovo. Obratimo sada pažnju na dve specifične stvari.
Prvo, slatkovodni kičmenjaci su evoluirali bubrege specijalizovane za proizvodnju velikih količina veoma razređenog urina (da bi iz tela izbacili što više vode, i što manje soli), što zahteva aktivno pumpanje soli prema unutrašnjosti tela (slatkovodni beskičmenjaci koriste sličan sistem, ali kroz drugačije sisteme organa).
Drugo, velika koncentracija soli u unutrašnjosti tela znači da se ta so brže gubi, i da neželjena voda većom brzinom ulazi u telo. Iz ovog razloga, slatkovodni kičmenjaci su evoluirali način života koji zahteva minimum soli – njihova ćelijska koncentracija soli je otprilike jedna trećina koncentracije koju nalazimo u morskoj vodi.
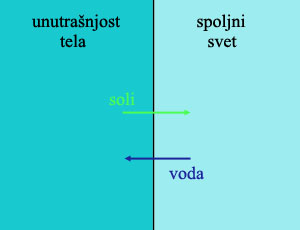
Ovo nas dovodi do ključne stvari: kičmenjaci sa lobanjom (takozvani kranijati) su evoluirali u slatkoj vodi, što znači da su evoluirali ovaj sistem bubrega, i nižu količinu soli u ćelijama. Povrh toga, nakon što su se ove osobine razvojno ustalile, one nisu mogle prosto da se uklone – što znači da su ove osobine zatim nasledili svi njihovi potomci. Time dolazimo do veoma zanimljive situacije koju vidimo danas.
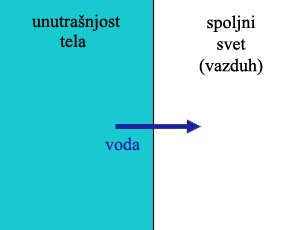
Naime, nakon što su se kičmenjaci iz slatke vode vratili u more, naišli su na suprotan problem. Odjednom, njihove ćelije su sadržale mnogo manje soli nego okolna voda. Znači, so stalno ulazi u njihovo telo, a pritom neprestano gube vodu! Da bi kompenzovale za ovo, morski kičmenjaci neprestano piju, i moraju da iz tela izbace što manju količinu što koncentrisanije tečnosti. Za ovo, potrebni su im sistemi koji pumpaju so iz tela napolje, obrnuto od slatkovodnih kičmenjaka. No, ovde nalazimo prvu zanimljivu stvar: njihovi bubrezi nisu napravljeni da rade ono što treba. Umesto toga, oni su nasledili bubrege slatkovodnih kičmenjaka, koji i dalje pumpaju so unutra i vodu napolje, i koji time proizvode urin koji je manje koncentrisan od okolne morske vode. Da bi kompenzovali za gubitak vode, ove životinje aktivno troše energiju pumpajući vodu nazad u telo kroz škrge, i pumpajući so napolje. Sve današnje morske ribe pate od ovog problema!
Povrh ovoga, imamo dodatnu komplikaciju sa tetrapodima (vodozemci, reptili, ptice, sisari…). Sve ove životinje su takođe nasledile unutrašnju koncentraciju soli od svojih slatkovodnih predaka, tj. otprilike 1/3 koncentracije prisutne u morskoj vodi. One takođe (uključujući tu i čoveka) i dalje nose isti osnovni dizajn: bubrege koji pumpaju razređenu vodu iz tela, i troše energiju da zadrže što više soli u telu. Naša fiziologija je napravljena za život u slatkoj vodi.
Pošto ove životinje ne dišu kroz škrge, one nemaju isti problem kao slatkovodne ribe, i ne moraju da se brinu oko suvišne vode koja ulazi u telo, ili dragocene soli koja izlazi. Ali one imaju jedan drugi problem: kroz disanje i kroz izbacivanje retkog urina, one gube velike količine vode, a zadržavaju velike količine soli u telu. Otud, svi tetrapodi moraju neprestano da nekako unose vodu u organizam, i moraju da nekako izbace višak soli.
Vodozemci nikada nisu prevazišli ovaj problem, i do dan danas žive isključivo u slatkovodnoj sredini (postoji samo jedan izuzetak, koji je evoluirao veoma specifične žlezde, i na osnovu toga može da živi u delimično slanoj vodi). Reptili i praktično sve ptice takođe ni dan danas ne mogu da proizvedu koncentrisani urin, već koriste posebne žlezde u očima i nosu, kroz koje izbacuju suvišnu so iz krvi (ove žlezde takođe nisu potpuno nova stvar, već su adaptacije prethodno postojećih suznih i pljuvačnih žlezda).
Sisari su evoluirali drugačije rešenje, okrećući nasleđene bubrežne pumpe u specijalnu petlju (takozvana Henleova petlja), u kojoj se naizmeničnim pumpanjem i ispumpavanjem soli proizvodi koncentrisani urin. Ptice-pevačice (Passeriformes) su nezavisno evoluirale strukturalno i razvojno drugačiji pristup koji dovodi do istog konačnog rešenja.
Ova potpuno nepotrebna dvostruka petlja je primer štetnog evolucionog nasleđa: deo strukture tela koje se ne možemo otarasiti, i koja čini naše bubrege mnogo osetljivijim nego što bi inače bili.
Primer: Ljudsko telo. Ljudsko telo sadrži mnogobrojne zaostatke evolucije.
Recimo, Kičma se završava nizom stopljenih repnih vertebri, sve sa neupotrebljivim mišićem za pokretanje tog nepostojećeg repa (extensor coccygis). Ušne školjke su opremljene sa po tri mišića za pokretanje ušiju – i sva tri mišića su i dalje prokrvljena i imaju svoje nerve, iako su potpuno nemoćni da pokrenu ljudsko uho. Ljudska vilica je postala smanjena kroz evoluciju, ali geni za broj zuba se još nisu adaptirali, zbog čega većina nas ima bolne, urasle umnjake koje moramo da vadimo (ili da se sa njima patimo celog života).
Evolucija disajnog puta od jednjaka čini da svi tetrapodi imaju rizik da im hrana zapadne u disajne kanale; ali ljudska evolucija govora je dovela do produbljenja larinksa, zbog čega smo mi posebno osetljivi na ovaj problem; na hiljade ljudi, većinom dece, umiru svake godine kroz davljenje hranom.
Možemo ovako nastaviti još dugo, ali za kraj ovog teksta, koncentrišimo se samo na posledice jedne nedavne adaptacije koja svima nama mnogo znači: uspravan položaj ljudskog tela. Isto kao i sa svim drugim osobinama svih drugih vrsta, naše telo nije napravljeno da bude uspravno, već je adaptirano iz prethodnog pognutog položaja kakav i dalje vidimo kod današnjih šimpanzi. Ovo se može videti na mnogo različitih osobina (i još jasnije u razvojnoj sekvenci), iz kojih ćemo se koncetrisati na tri:
1. Ljudska leđa. U fosilima ljudskih predaka možemo da vidimo kako se kičma postepeno ispravljala, i kako se karlica razvijala u skladu sa sve uspravnijim i uspravnijim položajem tela. No, isto kao sa “Baha Buba” primerom iz prethodnih poglavlja, adaptacija postojećeg oblika nikada ne može da bude onoliko dobra kao proizvodnja novog. Pošto su naša leđa samo adaptacija leđa prilagođenih za četvoronožni hod, ona nose mnoge zaostatke iz tog perioda, među kojima je i pelvis nagnut unapred. Zbog ovakvih stvari, ona su inherentno slaba i loše prenose sile koje nastaju u toku uspravnog hoda. Rezultat je da preko trećine čovečanstva pati od bolova u leđima, i da jedno nepažljivo dizanje tereta veće težine može dovesti do ozbiljne povrede leđa. Dodatni rezultat je smanjena fizička snaga; odrasli šimpanza može lako odraslom čoveku da iščupa ruku iz ramena, dok čovek tako nešto nikada ne bi mogao da učini šimpanzi. Uspravni hod koji nam je omogućio slobodne šake i korišćenje alatki ima svoju evolucionu cenu.
2. Sinusi. Delimično zbog smanjenja ljudskog lica, i delimično zbog rasta mozga, ljudski nosni sinusi su posebno tesno stisnuti. Ovo stiska kanale kojima se oni prazne, i usporava pražnjenje nagomilanih otpadaka. Povrh toga, kanal sfenoidnog sinusa je zbog ugnježdene hijerarhije razvoja okrenut – uvis. Zbog ovakvog slabog pražnjenja, bakterije se slobodno množe u sinusnoj sluzi i tečnosti, zbog čega mnogi od nas pate od čestih sinusnih infekcija i bolesti.
3. Trudnoća. Možda vizuelno najbolji primer efekata uspravljanja na ljudsku fiziologiju je raspored stomaka žene. Pogledajmo presek kroz donji abdomen ženskog tela:
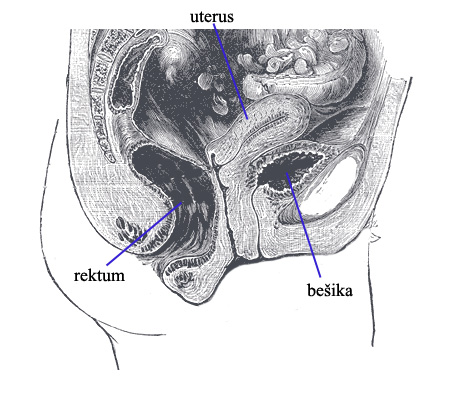
Raspored i građa organa je nasleđena kroz ugnježdenu hijerarhiju. No, posledice su velike. U kom pravcu leži težina deteta u materici tokom trudnoće? Pa, uporedimo: na donjoj slici označimo veličinu materice u toku trudnoće plavom isprekidanom linijom. Kod četvoronožnih životinja, kojima je stomak paralelan sa zemljom, gravitacija vuče dete u pravcu zelene strelice. Kod uspravnog čoveka, težina ide prema karlici (crvene strelice):
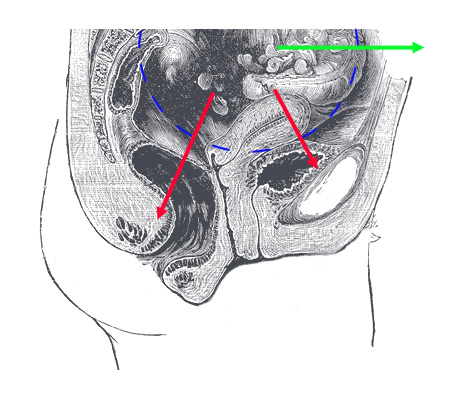
Rezultat je da materica u toku cele trudnoće “sedi” na bešici i rektumu, sa posledicama koje su poznate svim trudnicama (i njihovim muževima). Zbog ovakvog rasporeda organa, trudnoća često proizvodi i dugotrajne posledice po zdravlje žene. No, ovaj raspored se ustalio kroz stotine miliona godina evolucije, i zadnjih par miliona godina uspravnog hoda nisu dovoljni da ga značajno promene…
Ovo su samo neki lakše objašnjivi primeri tragova i nuzefekata evolucije. Prava i potpuna slika postaje vidljiva tek sa stanovišta biohemije i genetike, kada se na molekularnom i ćelijskom nivou posmatra razvoj organizama, i kada se jasno vidi kako nasleđene osobine ograničavaju razvoj budućih. No, nadamo se da su prethodni primeri dovoljni da se stekne makar opšta ideja ove linije dokaza.
Za kraj, u sledećem, zadnjem poglavlju, pogledaćemo šta nam sva prethodna govore o prirodi i razvoju živog sveta na našoj planeti.
[prethodno poglavlje][nazad na vrh strane][sledeće poglavlje]