
- Šta je uopšte evolucija
- Evolucija danas: mutacije i nastanak informacija
- Evolucija danas: selekcija i širenje informacija kroz populaciju
- Evolucija danas: prirodna selekcija, geni i populacije
- Teorija evolucije života na Zemlji
- Dokazi: Evoluciona istorija i ugnježdena hijerarhija
- Dokazi: Genetika, genomika i tragovi evolucije
- Dokazi: Fosilni zapis i razvoj života
- Dokazi: Razvojni i fiziološki tragovi
- Zakoni prirode ili slučajna konfluenca dokaza?
Kao što smo videli u prethodnom poglavlju, nasleđe sa modifikacijom proizvodu ugnježdenu hijerarhiju. Ova hijerarhija je najbolje vidljiva na genetskom nivou. Svaka pojedinačna promena i mutacija se nasleđuje, i te promene se mogu pratiti kroz stablo nasleđa.
Opet, ovde nije reč o sličnosti – recimo, genom čoveka nije prosto sličan genomu šimpanze, već su oba genoma kopije sa modifikacijama jednog istog početnog.
Od ključne važnosti je činjenica da se genomi kopiraju u celini. Vrsta mutacije poznata kao delecija povremeno “briše” delove genoma, ali ovo se događa veoma sporom brzinom. Iz ovog razloga, genomi u sebi sadrže ostatke prethodne evolucije organizma. Razjasnimo ovo u tri koraka:
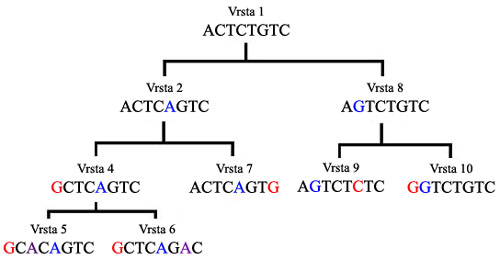
– Najjednostavniji nivo je praćenje promena. U prethodnom pogavlju smo upotrebili primer ugnježdene hijerarhije genoma psa:
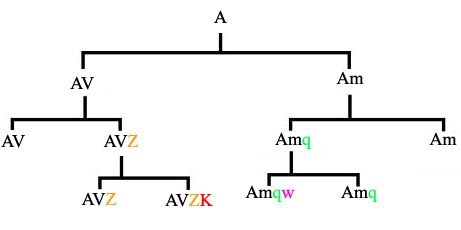
Sa slike je očigledno da se sve promene prenose na buduće rase pasa kroz veštačku selekciju. U prirodi, promene se prenose na podvrste pa zatim na vrste, kroz prirodnu selekciju. U gornjoj shemi, vrsta koja nosi gene AVZK je nastala od vrste koja nosi gene AVZ – ili, drugim rečima, njeni geni su potpuna kopija svih gena AVZ uz naknadnu modifikaciju K.
– U drugom poglavlju, videli smo da je duplikacija praćena modifikacijama jedan od glavnih načina na koji nastaju novi geni. Slično prethodnom, ovo znači da možemo nastanak novih gena da pratimo korak po korak. Recimo, možemo da vidimo u jednom organizmu gen X, dok u drugom organizmu vidimo isti taj gen X, plus gen koji je skoro identičan genu X, samo sa dodatnim izmenama – rezultat kopiranja i modifikacije, istog procesa koga danas vidimo u prirodi.
Velika većina gena spada u ovu kategoriju. Recimo, kontrolni geni poznati kao kinaze, su se razvili kroz duplikaciju i modifikaciju od jednog pretka, što nam omogućava da rekonstruišemo njihovo stablo nastanka:
Kao što možemo videti, ovo stablo je “malčice” komplikovanije od prethodnih koje smo koristili za ilustraciju. No, ono je lepa ilustracija koliko osnovne osobine gena ostaju očuvane kroz evolutivni razvoj, i kroz ponovne i ponovne procese duplikacije sa modifikacijama.
– U primeru pećinskih riba, videli smo da uklanjanje selektivnog pritiska dovodi do gubitka osobine. Ali gubitak osobine jako retko znači da je gen za tu osobinu obrisan iz genoma. Naprotiv, ono što se događa je da se u okviru gena dese mutacije koje taj gen “pokvare” i deaktiviraju.
No, ovaj deaktivirani gen se i dalje prenosi kroz generacije, sa tim mutacijama koje su ga uništile. Ako se vrsta podeli na dve, obe novonastale vrste će i dalje imati u svom genomu mutirani ostatak početnog gena. Štaviše, ovaj ostatak će i dalje nakupljati mutacije, omogućavajući nam da napravimo još jedno nezavisno stablo nasleđa na osnovu ovih razlika.
Sva ova stabla prate isto osnovno stablo nasleđa koje možemo rekonstruisati na osnovu morfoloških osobina, na osnovu fosila (kao što ćemo videti u sledećem poglavlju), na osnovu biohemijskih sposobnosti, na osnovu fiziologije…
Ostaje pitanje kako znamo da je ovo zaista produkt evolucije, a ne produkt nekakvog namernog mešanja ili planiranog dizajna? Za ovo postoji više razloga, od kojih ćemo se koncentrisati na četiri:
- Neuklapanje verovatnoća. Ako se genetski kod namerno menja, on se može menjati na bilo kakav način: bilo koje “slovo” se može promeniti, prepisati ili izbrisati. Prirodni procesi koje posmatramo u svetu danas, međutim, proizvode određene vrste promena mnogo češće nego druge. Otud, kada pogledamo kojim promenama su današnji genomi nastali iz prethodnih, možemo da vidimo da su oni menjani prirodnim procesima, a ne veštačkom intervencijom.
- Nasleđe degenerisanih osobina. Kao što ćemo videti u sledećim primerima, geni koji su potpuno nefunkcionalni se i dalje nasleđuju.
- Nedostatak prebacivanja gena. Svi geni koji se pojavljuju unutar neke vrste u ovoj hijerarhiji su uvek kopije gena iz prethodnih koraka ove hijerarhije. Ovo znači da se “topla voda uvek ponovo otkriva” umesto da se prebace prethodno postojeći oblici. Recimo, geni koji zečevima omogućavaju preživarski način života su nastali kopiranjem i modifikacijom gena koji su bili prisutni u precima zečeva – umesto da se prosto prebace potpuno funkcionalni geni koji istu sposobnost omogućavaju kopitarima.
- Poklapanje stabala. Stabla nasleđa razvijena na različite načine se poklapaju jedna sa drugim. Genetsko stablo nasleđa se poklapa sa morfološkim, fiziološkim, genetskim, sa nasleđem degenerisanih osobina, itd. Verovatnoća da je ovo slučajno tako je nezamislivo mala.
Uzmimo sada nekoliko konkretnih primera iz moderne genetike. Počnimo sa jednim primerom koji ilustruje verovatnoće o kojima govorimo ovde.
Primer: Matematičko praćenje stabla života. Geni se sastoje od uređenog niza sa dužinom od više stotina, hiljada ili čak miliona DNK “slova”. Recimo, evo kako izgleda sekvenca jednog veoma kratkog gena (CFP gen, originalno iz jedne vrste meduza):
GGTACCAATGTCCAAAGGTGAAGAACTTTTCACTGGAGTGGTGCCTATCTTGGTTGAGCTTGACGGTGAT GTGAACGGTCACAAATTCTCTGTAAGTGGTGAAGGAGAGGGAGACGCTACCTACGGCAAGTTAACGCTGA AATTTATATGCACTACGGGAAAGCTGCCTGTACCGTGGCCTACACTGGTTACCACTCTGACTTGGGGAGT ACAATGCTTCGCCCGCTATCCGGACCACATGAAACGCCATGACTTCTTTAAATCAGCTATGCCAGAAGGA TACGTGCAAGAACGAACCATTTTCTTCAAAGATGATGGTAATTATAAAACAAGGGCGGAAGTTAAATTCG AAGGAGACACGCTCGTAAACAGAATCGAACTTAAGGGTATCGACTTCAAGGAGGATGGCAACATTCTTGG ACACAAACTGGAGTACAACGCCATTTCCGATAATGTTTACATTACTGCTGATAAACAGAAGAACGGAATT AAGGCGAATTTTAAAATCAGACATAACATTGAAGATGGTGGAGTGCAATTGGCTGATCACTACCAGCAAA ATACTCCTATCGGAGACGGCCCTGTGTTGCTTCCTGACAACCACTACTTAAGTACGCAATCAGCTTTATC CAAAGATCCCAATGAAAAGCGAGATCACATGGTGCTGCTCGAATTTGTTACTGCTGCTGGTATTACACAC GGAATGGATGAGCTGTATAAATAAGAATTC
Ovaj gen je dugačak 730 “slova”, ili tačnije, baznih parova. Postoje geni koji su dugački više hiljada ili više miliona baznih parova. Uzmimo da sada, danas, u prirodi vidimo kako je u organizmu nastao još jedan gen, koji izgleda ovako:
GGTACCAATGTCCAAAGGTGAAGAACTTTTCACTGGAGTGGTGCCTATCTTGGTTGAGCTTGACGGTGAT GTGAACGGTCACAAATTCTCTGTAAGTGGTGAAGGAGAGGGAGACGCTACCTACGGCAAGTTAACGCTGA AATTTATATGCACTACGGGAAAGCTGCCTGTACCGTGGCCTACACTGGTTACCACTCTGACTTGGGGAGT ACAATGCTTCGCCCGCTATCCGGACCACATGAAACGCCATGACTTCTTTAAATCAGCTATGCCAGAAGGA TACGTGCAAGAACGAACCATTTTCTTCAAAGATGATGGTAATTATAAAACAAGGGCGGAAGTTAAATTCG AAGTAATAGGCTCGATCGAGAGACACGCTCGTAAACAGAATCGAACTTAAGGGTATCGACTTCAAGGAGG ATGGCAACATTCTTGG
Ovaj gen se sastoji iz stotina baznih parova koji se potpuno poklapaju sa prethodnim genom, uz jednu grupu dodatnih “slova” (označenih crvenom bojom). Ovde imamo dva izbora. Ili je ovaj gen nastao potpuno slučajno, ili je nastao kroz kopiranje jednog dela prethodnog gena uz dodatno ubacivanje jednog dodatnog dela.
Možemo izračunati verovatnoću za oba izbora. Kolika je verovatnoća da se pojavi novi gen koji se slučajno poklapa sa već postojećim? Ova verovatnoća je veoma mala, toliko mala da je krajnje neverovatno da se tako nešto može desiti čak i kroz milijarde godina mutacija. Sa druge strane, koja je verovatnoća za kopiranje i ubacivanje dodatnog koda? Ako je ovo kod iz prosečne populacije bakterija, šanse su da će se ovako nešto desiti jednom svakih nekoliko dana; kod kičmenjaka ili ljudi, desiće se jednom u svakih nekoliko decenija.
Dakle, jedna opcija je krajnje neverovatna, dok je druga opcija sasvim moguća.
No, ovo je računica za jedan pojedinačni gen. Mi danas imamo sekvence celih genoma – milijarde DNK “slova”, u stotinama i hiljadama vrsta. Ove sekvence se mogu takođe uklopiti u stablo hijerarhije, i mogu se izračunati verovatnoće da se naši podaci slučajno uklapaju u ovu hijerarhiju.
Pogledajmo kako izgleda jedno filogenetsko stablo izračunato na osnovu celokupnih genoma par stotina organizama:
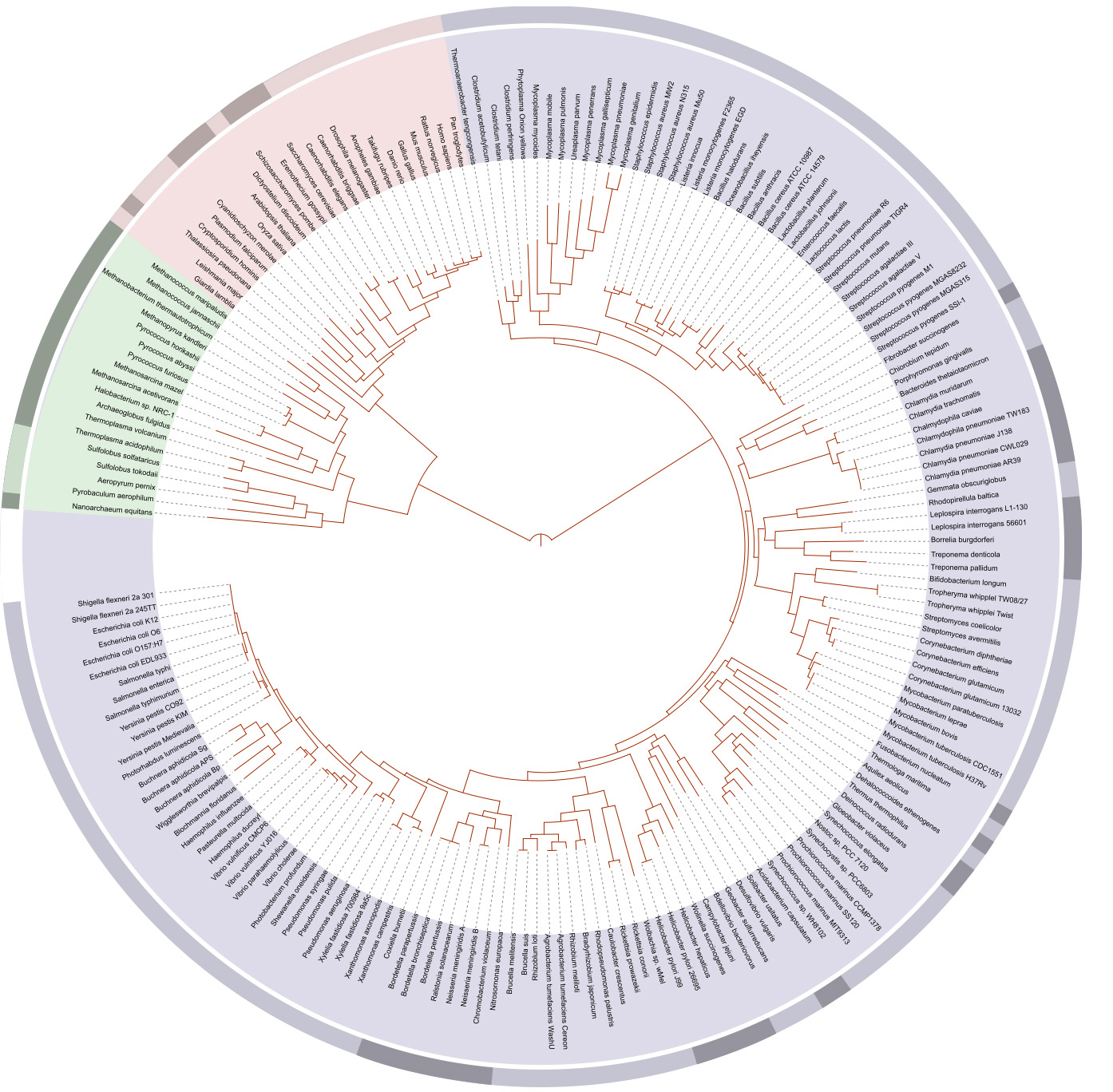
(Kliknite na sliku za veću verziju.)
Mi možemo sada izračunati kolika je verovatnoća da se ovi geni slučajno uklapaju u stablo nasleđa. Ta verovatnoća je jednaka sledećoj cifri:
1 u 250000000000000000000000000000000000000000000000000000000000000000000000000000
Ovo je toliko mala verovatnoća, da ne možemo naći realno poređenje za nju u stvarnom životu. Recimo, verovatnoća da će čitalac ovog teksta slučajno dobiti sve svetske lutrije svaki put od sada do kraja života je neuporedivo veća od šanse da se slučajno desi ovakvo poklapanje stabala nasleđa.
Okrenimo se pitanju prenosa nefunkcionalnih osobina, na jednom primeru koji uključuje i ljudski genom.
Primer: Zašto je ljudima potreban vitamin C? Vitamin C (takođe poznat kao askorbinska kiselina) je supstanca potrebna u mnogim biohemijskim reakcijama, zbog čega se ona proizvodi u ćelijama praktično svih biljaka i životinja.
Nekoliko vrsta životinja je, međutim, izuzetak od ovog pravila. Ako se životinja hrani velikim količinama voća, ona u telo unosi velike količine vitamina C. Samim tim, ovo uklanja selektivni pritisak koji održava postojanje gena za njegovu proizvodnju.
Proces gubitka osobina nakon nestanka selektivnog pritiska smo već ilustrovali na primeru vida pećinskih riba; ali pošto je poenta veoma važna, vredi opisati proces još jednom. Ako tigar u divljini okoti deset tigrića, i jedan od tih deset nosi mutaciju koja onemogućava proizvodnju vitamina C u telu – taj tigrić će ubrzo uginuti od skorbuta. Sa druge strane, ako neka životinja jede prevashodno voće, i ako ta životinja ostavi potomka koji doživi istu ovu mutaciju, taj potomak će preživeti sasvim normalno, pošto iz ishrane dobija daleko više vitamina C nego što mu je potrebno. Mutacija koja onesposobljava proizvodnju vitamina C se zatim prenosi dalje na potomke ove jedinke.
Zbog ovoga, mnoge vrste koje se hrane voćem izgube sposobnost proizvodnje vitamina C. Ovaj gubitak se onda prenosi i na dalje generacije, pa i na vrste koje nastanu od ove početne, čak i ako ta vrsta promeni ishranu.
Upravo ovo se desilo i sa primatima. Predak današnjih tarsijera, majmuna, hominida, i ljudi je doživeo mutaciju koja je deaktivirala takozvani GULOP gen (ovaj gen proizvodi enzim L-gulonolakton oksidazu, neophodan za proizvodnju vitamina C). Ovaj deaktivirani gen se onda prenosio dalje kroz kopiranje genoma sve do danas. Danas možemo da posmatramo kako su se i kada određene promene na ovom genu događale, i kako su bile prenošene sa predaka na potomke.
U okviru ovog teksta, mi ne možemo prikazati celo stablo nasleđa ovog gena, pošto bi smo dobili daleko previše kompleksnu sliku (sličnu onoj gornjoj koja je data za kinaze). Ali možemo ovo prikazati uprošćeno: označićemo GULOP gen prostom plavom linijom, i onda ćemo na njemu označavati samo velike promene, i to samo one koje su se prenele do čoveka danas. Dakle, shema koja sledi prikazuje razvoj ovog gena sa ljudske tačke gledišta:
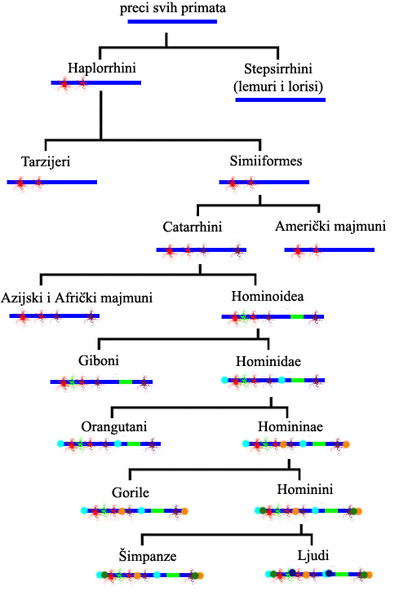
Dakle, preci svih primata su nosili u sebi aktivan i normalan GULOP gen. Ova grupa se podelila na dve. Sa jedne strane, odvojili su se Stepsirhini, preci današnjih lemura i lorisa, koji su nasledili normalan gen i imaju normalnu sposobnost proizvodnje vitamina C. Sa druge strane su se odvojila grupa Haplorhini, koji su se orjentisali na ishranu bogatu voćem (ovo vidimo iz adaptacija njihovog zubnog aparata, adaptacija tela, i izotopskih studija njihovih ostataka). Negde u toku ovog perioda, ovaj GULOP je doživeo mutacije koje su ga deaktivirale. No, deaktivirana, mutirana kopija gena je ostala u genomu Haplorina.
Nešto kasnije, jedna grupa Haplorina koja je u svom genomu nosila tu mutaciju se podelila na više, od kojih su nastali tarzijeri, američki majmuni, i grupa poznata kao Katarini. Sve ove tri grupe su nasledile iste one početne destruktivne mutacije, i nijedna od ove tri grupe nema sposobnost da sama proizvodi svoj vitamin C. Sve grupe su se zatim dalje delile na nove vrste, i nakupljale su nove mutacije u GULOP genu, ali mi ćemo se koncentrisati na Katarine, kod kojih je GULOP gen doživeo još nekoliko specifičnih mutacija, dodatnih oštećenja na već pokvarenoj sekvenci.
Par miliona godina kasnije, Katarini su se i dalje podelili: sa jedne strane su se odvojili preci današnjih afričkih i azijskih majmuna, a sa druge su se odvojili Hominoidi. Opet, obe grupe su nasledile četiri mutacije od Katarina, pa su na njih dodali svoje dodatne mutacije. Kod Hominoida, u sekvencu GULOP gena se ubacilo parče virusne DNK poznato kao retrotranspozon. Svi naslednici Hominoida su nasledili GULOP gen sa ovom ubačenom sekvencom, povrh svih ostalih već pomenutih mutacija.
Nadamo se da je shema nasleđa ovde jasna. Hominoidi su se podelili na gibone i Hominide, Hominidi na orangutane i Homininae…itd, sve do zadnje podele u kojoj su se razdvojili preci današnjih šimpanzi i današnjeg čoveka. U svakoj podeli, svaka strana je na prethodno nasleđene mutacije dodavala svoje specifične, gradeći stablo nasleđa korak po korak. Prateći zajedničke mutacije, možemo videti ugnježdenu hijerarhiju porekla.
Ovakvi deaktivirani geni se zovu “pseudogeni”, i ima ih u genomima svih organizama. U ljudskom genomu ih ima na stotine – između ostalog, recimo, preko šest stotina deaktiviranih gena za mirisne receptore (koji su mutirali kako je u istoriji ljudskih predaka čulo mirisa postajalo sve manje bitno). Svi ovi geni se mogu koristiti za praćenje ugnježdene hijerarhije nasleđa.
Primer: Retrovirusni ostaci. Većina virusa deluje tako što u ćeliju ubaci svoj genetski materijal, koji onda preuzme kontrolu nad ćelijom. Umesto svojih normalnih procesa, ćelija počne da proizvodi nove kopije virusa. Na kraju, ćelija umire i raspadne se, oslobađajući nove viruse, koji sada mogu da napadnu druge ćelije.
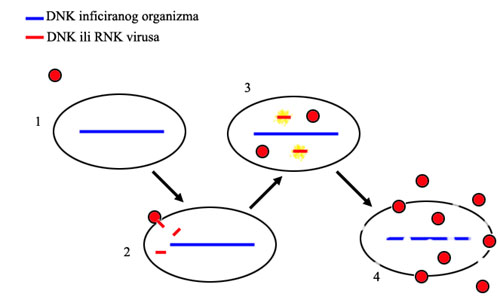
Tok virusne infekcije: 1. virus se približi ćeliji, 2. virus se spoji sa ćelijom i u nju ubaci svoj genetski materijal, 3. geni virusa postanu aktivni i ćelija na osnovu njih proizvede još virusa, 4. ćelija umire i raspada se, oslobađajući gomilu novih virusnih čestica.
Retrovirusi idu korak dalje: oni ne samo što ubace svoj genetski materijal u ćeliju, već ga zatim i integrišu u genom “domaćina.” Rezultat je da ćelija ne može da razlikuje virusne gene od svojih sopstvenih. Virus ne mora čak ni da bude aktivan, može da se ućuti i da prosto čeka, što mu omogućava da prebrodi imunu reakciju unutar tela. Onda, u nekom budućem trenutku, geni se aktiviraju, i preuzmu kontrolu nad ćelijom; ćelija zatim umre proizvodeći gomilu novih virusa. Najpoznatiji retrovirus danas je HIV, virus koji uzrokuje sidu.
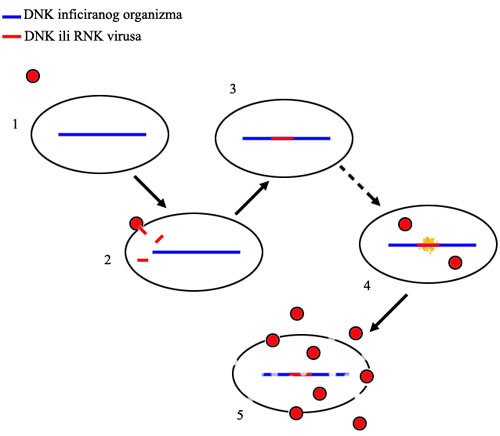
Tok retrovirusne infekcije: 1. retrovirus se približi ćeliji, 2. retrovirus se spoji sa ćelijom i u nju ubaci svoj genetski materijal, 3. geni virusa se utope (integrišu) u genom organizma, i tu mogu da ostanu mesecima ili godinama bez aktivnosti, 4. geni postanu aktivni i ćelija na osnovu njih proizvede još virusa, 5. ćelija umire i raspada se, oslobađajući gomilu novih virusnih čestica.
U dodatnom tekstu je objašnjeno kako ovi virusni napadi mogu postati izvor novih gena. No, ovi napadi takođe ostavljaju jasne tragove u vidu ostataka virusnih gena integrisanih u genom organizma.
Nekodirajuća DNK
Koliki deo ljudskog genoma čine geni, a koliki druge stvari? Da li postoji neupotrebljiva DNK, i koliki deo genoma ona čini? Pogledajte dopunski tekst na ovu temu.
Ovo se događa na veoma prepoznatljiv način: integracija virusnih gena, čak i ako je neuspešna, ostavlja trag u vidu dugog genetskog niza. Taj niz je, u neku ruku, genetski ožiljak: trag virusnog napada na jedan određeni organizam. Ovaj “ožiljak” je pritom veoma specifičan: pošto se sastoji od više stotina ili hiljada genetskih slova, efektivno je nemoguće da dva organizma slučajno steknu isti ožiljak na istom mestu.
Tako, recimo, svi ljudi na svetu nose u svom genomu nekoliko tragova napada virusa poznatog kao HERV-K. Ovaj virus je napao pretke ljudi pre par stotina hiljada godina, i tragove ovog napada svi nosimo u svojim genima do dana današnjeg.
Ali ovi retrovirusni tragovi, isto kao i mutacije ili pseudogeni, mogu da se nasleđuju i kroz stablo grananja vrsta! Mada ovako možemo analizirati praktično svaku granu života (jako lep primer su retrovirusni tragovi u genomu mačaka, na primer), koncentrišimo se ponovo na ljudske pretke, i samo na ostatke jedne već pomenute familije virusa, HERV. Od ukupno četrnaest (14) jedinstvenih tragova napada ovakvih virusa u genomu čoveka, identične tragove na identičnom mestu nalazimo:
- na jedanaest (11) mesta kod šimpanzi i gorila
- na devet (9) mesta kod orangutana
- na sedam (7) mesta kod gibona
- na četiri (4) mesta kod Afričkih i Azijskih majmuna
- na dva (2) mesta kod Američkih majmuna
Sad, podsetimo se: verovatnoća da dva retrovirusa napadnu dva različita genoma, ali da se udenu na tačno isto mesto i ostave tačno isti trag je neverovatno mala. Šansa da se dva virusa slučajno ubace na isto mesto i ostave isti trag u različitim genomima je još neuporedivo manja – otprilike jednaka verovatnoći da molekuli koji čine planetu Zemlju odjednom slučajno zavibriraju na takav način da se čitava planeta razleti na sastavne atome.
Jedini realan način na koji ovakvi tragovi mogu nastati je nasleđe: virus napadne genom jednog organizma, ostavi trag, i svi potomci tog organizma nasleđuju isti “ožiljak” na istom mestu u genomu. Stablo nasleđa koje dobijamo na ovaj način se savršeno uklapa stablom koje smo ranije dobili kroz poređenje celih genoma, ili sa stablom koje smo dobili poređenjem pseudogena, ili koje možemo dobiti na razne druge načine:
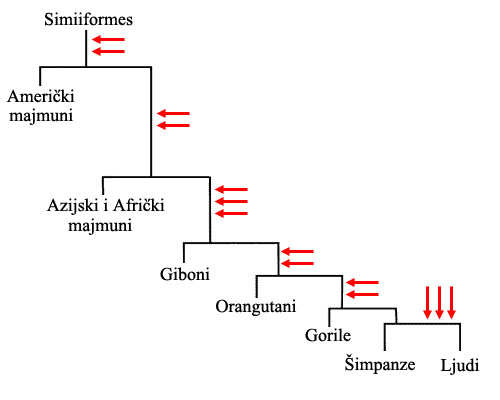
Na gornjoj shemi, svaka crvena strelica označava jedan virusni napad. Svaki takav napad je ostavio jedinstven trag u genomu inficiranog organizma, trag koga su nasledili njegovi ili njeni potomci…
Ovo je jedan mali izbor genetskih primera evolucije; veliki deo zahteva mnogo dublje ulaženje u genetiku. No, gornji primeri bi trebalo da budu dovoljna ilustracija razloga zbog kojih genetski podaci podržavaju evolucioni model. Pređimo sada na sledeće poglavlje, u kome ćemo pogledati najdirektniji primer evolucione istorije života: prelazne fosile.
[prethodno poglavlje][nazad na vrh strane][sledeće poglavlje]