
- Šta je uopšte evolucija
- Evolucija danas: mutacije i nastanak informacija
- Evolucija danas: selekcija i širenje informacija kroz populaciju
- Evolucija danas: prirodna selekcija, geni i populacije
- Teorija evolucije života na Zemlji
- Dokazi: Evoluciona istorija i ugnježdena hijerarhija
- Dokazi: Genetika, genomika i tragovi evolucije
- Dokazi: Fosilni zapis i razvoj života
- Dokazi: Razvojni i fiziološki tragovi
- Zakoni prirode ili slučajna konfluenca dokaza?
U većini slučajeva, ubrzo nakon smrti, ostaci biljaka i životinja bivaju razoreni kroz procese truljenja, ostavljajući u najboljem slučaju samo nekoliko malih komadića najtvrđih kostiju. No, povremeno, u retkim okolnostima, ovi ostaci se nađu u uslovima u kojima se tkivo mineralizuje, što nam ostavlja okamenjeni trag životinje, njen fosil. Najčešće se fosilizuju oni delovi životinje koji se najsporije raspadaju (i otud imaju najviše vremena za mineralizaciju): zubi, vilice i ostale kosti. Zbog ovoga, skoro svi fosili su fosili životinjskih kostura. Skup svih fosila nađenih u sedimentnim slojevima u zemlji se naziva fosilni zapis, i on predstavlja fizički trag istorije života na našoj planeti.
Naravno, životinja se ne može fosilizovati pre nego što je nastala. Otud, fosilni zapis nam može poslužiti kao još jedan dodatni način kojim možemo nezavisno proveriti teoriju evolucije. Ako se u fosilnom zapisu životinje pojavljuju bez reda i poretka, sve odjednom, to bi svakako značilo da je teorija evolucije ili netačna, ili bar nepotpuna. Međutim, ono što smo našli je upravo suprotno.
Fosilni zapis pokazuje ukupnu ugnježdenu hijerarhiju živih bića: fosili iz kasnijih slojeva su kopije sa modifikacijama fosilizovanih životinja iz prethodnih slojeva. Samim ovim, fosilni zapis je u stvari zapis evolucije, pokazujući nam kako su izgledale vrste u prošlosti, i kako su se one razvijale kroz evolutivne procese.
Ako fosile koji se uklapaju u jednu celovitu hijerarhiju nasleđa poređamo po starosti, dobijamo fosilnu sekvencu određene vrste. Lep primer jedne takve sekvence su danas izumrli organizmi zvani titanotere, donekle slični današnjim nosorozima. Na donjoj slici sa leve strane, možete videti sekvencu fosila lobanja titanotera i njihovih predaka, poređanu po starosti (raniji oblici su na dnu, noviji na vrhu; rekonstrukcije izgleda za neke od lobanja su date sa desne strane):
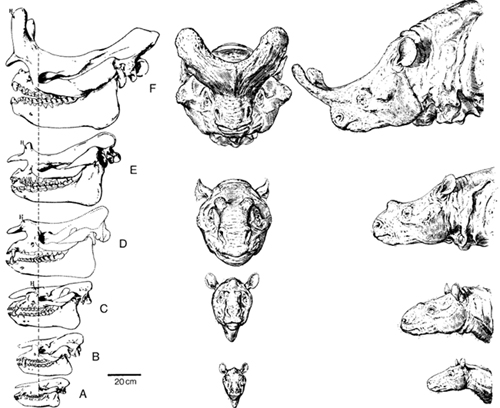
Adaptirano iz Steven M. Stanley, “Relative Growth of the Titanothere Horn: A New Approach to an Old Problem”, Evolution 1974, 28(3).
Ovo nam pokazuje kako se procesi evolucije nastavljaju i preko granica koje danas vidimo na primerima datim u prethodnim poglavljima. Recimo, razlika između lobanje A i C na slici je otprilike ekvivalentna spektru razlike koga danas vidimo kod pasa. Slično važi za lobanje od C do E, ili od D do F.
Direktnije rečeno, mi znamo, pošto smo to direktno videli, da procesi evolucije mogu proizvesti promenu od bića A do bića B; ili od bića B do bića C; ili od bića C do bića D…itd. Na žalost, ne možemo da posmatramo evoluciju stotinama hiljada ili milionima godina, i ne možemo direktno danas da vidimo koliko dugo takav proces može da ide, i kolika je dugoročna promena koju taj proces proizvodi.
Pomoću fosila, mi vidimo niz takvih koraka jedan iza drugog. Na osnovu toga, možemo direktno svojim očima da se uverimo da procesi evolucije ne idu samo jedan korak unapred, već da korak po korak mogu da značajno promene izgled i osobine vrste.
Ali ova gornja sekvenca je jedna mala sličica u okviru jedne grane kopitara – mali konjoliki kopitar se polako pretvorio u ogromnog nosrogolikog kopitara. Mada je ovo zanimljivo, većinu ljudi mnogo više zanimaju zaista veliki prelazi u evoluciji – kako su kopnene životinje evoluirale od riba, kako su nastale ptice, kako su nastali sisari…
U okviru ovog teksta, daćemo opšti pregled par specifičnih fosilnih nizova koji dobro ilustruju evoluciju. Čitaocima koji žele više detalja, predlažemo da nakon što završe čitanje “Uvoda u Evoluciju” svrate i do mnogo dužeg i detaljnijeg teksta na temu prelaznih fosila.
Primer: Evolucija kopnenih životinja. U okviru nastanka kopnenih životinja, prvi ključni korak se odigrao daleko pre nego što je bilo koja životinja stupila na suvo tlo. Molekularni, genetski, i morfološki dokazi pokazuju da su već rane koščate ribe posedovale takozvanu gastričku kesicu: proširenje sa strane jednjaka, u koje je riba mogla da “proguta” vazduh, što olakšava plivanje (ribe sa ovakvim kesicama mogu da plutaju, dok ribe bez njih – na primer, ajkule – moraju neprestano da ulažu energiju i plivaju, jer u suprotnom potonu na dno okeana).
Ova gastrička kesica je postala predmet selekcije. Kod riba u otvorenoj vodi, ona je postala ključna za plivanje, i razvila se u današnju vazdušnu bešiku.
Fosilni zapis
Tokom poslednjih decenija, ukupan broj prelaznih fosila je sa stotina prešao na hiljade, desetine i stotine hiljada, da bi do 2021-ve dostigao milione. U tekstovima na ovom sajtu, radi jednostavnosti, fokusiramo se na nekoliko najvažnijih. Za više detalja, možete pogledati paleontološke naučne žurnale, ili sajtove poput Palaeos.
Kod riba u plitkim i mutnim vodama siromašnim vazduhom, ova kesica se nalazila pod drugačijim selektivnim pritiskom. U ovakvoj vodi ribi je dostupno veoma malo kiseonika, a blato koje zaustavlja protok kroz škrge čini stvari još gorim. Otud, gastrička vrećica je kod njih postala prokrvljena, omogućavajući ribi da iz progutanog vazduha apsorbuje kiseonik. Takođe, svaka mutacija koja pomera otvor kesice bliže ustima je pozitivno selektovana, pošto smanjuje verovatnoću da progutani vazduh završi u želucu (gde kiseonik ne može efikasno da se apsorbuje).
Ovaj put razvoja je doveo do nastanka primitivnih ribljih pluća, koja možemo i dan danas da vidimo kod nekoliko ribljih grupa. Primer ovoga su ribe gar i amija među zrakoperkama, i riba dvodihalica među rezoperkama.

Kvinslendska dvodihalica, Neoceratodus forsteri, poseduje škrge, pluća, i koščata peraja. Photo: Tannin
Pre oko 400 miliona godina, kopnene biljke su konačno počele da stabilizuju priobalne ekosisteme, što je vodenim životinjama pružilo mogućnost novih načina života, i novih izvora hrane. Usled suše, ili čak prosto ciklusa plime i oseke, ove oblasti su često ostajale izolovane (u vidu muljevitih bara odvojenih od otvorene vode) i bile su siromašne kiseonikom. Otud, nije čudno što u fosilnim nalazištima iz priobalnih ekosistema ovog perioda često nalazimo upravo mnogo riba dvodihalica. Jedna grupa ovakvih riba, po imenu ripidistija (Rhipidistia), je posebno značajna.
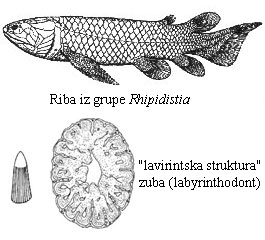
Ove ribe, kao i sve dvodihalice, su imale i pluća i škrge. Takođe, dok su “zraci” u perajima zrakoperki vezani direktno za osnovni skelet, kod ripidistijanskih riba i drugih rezoperki, ovi “zraci” su vezani za snažne kosti i mišiće. Konačno, imali su i osobinu koja je veoma jedinstvena za ovu grupu: njihovi zubi su imali veoma specifičnu “lavirintsku” strukturu.
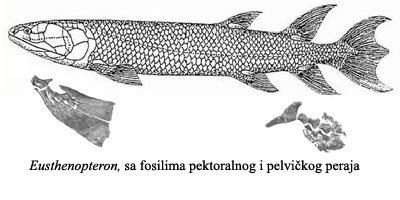
U fosilnom zapisu vidimo kako su se ripidistijanske ribe, izložene novim selektivnim pritiscima, brzo prilagodile svojoj novoj okolini. U okviru prilagođavanja na život u plićaku (umesto plivanja u dubokom moru), telo kasnijih ripidistija postaje spljošteno, oblik lobanje se menja, rep postaje prav. Zbog hroničnog nedostatka kiseonika, pluća postaju efikasnija i podeljena na dva dela. Konačno, kosti peraja postaju veće, snažnije, i mišići postaju snažniji. Ovakva peraja su skoro neupotrebljiva za plivanje kroz duboku vodu, ali su veoma korisna za razgrtanje gustog bilja u plićaku, i za puzanje kroz mulj. Primer jedne ovakve ribe je bio eustenopteron (Eusthenopteron):
Pritisci okoline su proizveli mnogo vrsta veoma bliskih Eusthenopteronu, ali koje su se još više odmakle od riba. Jeda lep primer ovoga je panderihtis (Panderichthys):

Autor: Nobu Tamura
Pošto više nije bilo selektivnog pritiska koji održava postojanje leđnih i analnih peraja, kod panderihtisa su ona zakržljala. Ostao je samo rep i kratka peraja na veoma snažnim i koščatim udovima. Povrh toga, kosti u okviru ovih udova su postale potpuno diferencirane, i to na veoma specifičan način: humerus, ulna i radius u prednjim udovima, i femur, tibija i fibula u zadnjim udovima. Takođe se pojavila čoana, prolaz u kosti koji povezuje nosnu šupljinu sa ustima, što omogućava disanje kroz nos.
Ovakve “puzeće vrste” se šire i granaju u toku sledećeg perioda vremena. Imamo mnogo prelaznih fosila iz ovog perioda, koji pokazuju razne vrste životinja sličnih panderihtisu (recimo Ventastega, ili Elginerpeton). Fosil koji nas posebno zanima je zvan tiktalik (Tiktaalik):
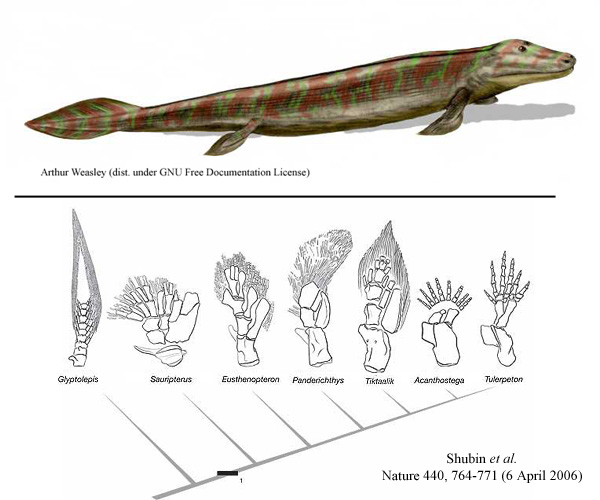
Kod tiktalika, vidimo još par malih promena (da ponovimo, svaki korak u evoluciji čine male, malecne promene koje se polako šire i nakupljaju). Najvažnije za nas su promene u strukturi udova: veza između kostiju koja je kod panderihtisa bila čvrsta, kod tiktalika postaje savitljiva (drugim rečima, nastaje zglob). Povrh toga, neki od “zraka” u okviru peraja postaju zadebljani, što omogućava životinji da se lakše odupre od tla, i samim tim da lakše puzi po zemlji; ova zadebljanja su u obliku niza falangijalnih kostiju (tj. kostiju prstiju).
Ova zadebljanja padaju pod selektivni pritisak za lakše kretanje kroz mulj i preko kratkih distanci na suvom tlu. Posle samo par miliona godina, vrste koje su nasledile ove osobine se šire svetom, toliko da neke od njih nalazimo u mnogim naslagama priobalnih područja u svetu. Jedan primer ovakve uspešne vrste je akantostega (Acanthostega):

Ova “žaboriba” je imala i pluća i škrge. Mada su kosti lobanje identične kasnijim vodozemačkim, oblik lobanje je još uvek riblji. Rep još uvek ima riblje karakteristike, sa perajem na leđnoj strani. Veza između lobanje i ostatka tela je riblja. Zubi i dalje imaju lavirintsku strukturu, nasleđenu od ripidistijanskih riba. I dalje postoji lateralna linija, organ koga ribe koriste da osete vibracije u vodi (ali koga vodozemci nemaju). Kada se sve uzme u obzir, nemoguće je reći da li se radi o ribi ili o vodozemcu – Acanthostega je savršena prelazna forma između dve klase živih bića.
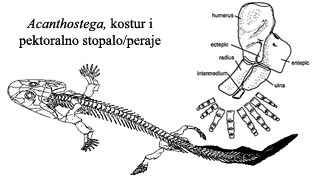
Kod akantostege, raspored i broj kostiju u “rukama” i “nogama” je identičan onome koga vidimo kod svih kopnenih kičmenjaka (guštera, ptica, sisara…). Pozicija stopala je, međutim, još uvek prilagođena vodenom životu, i nepogodna za hodanje po suvom tlu. Ovaj razvoj vidimo kod jedne srodne vrste iz istog perioda, ihtiostege (Ichtyostega):
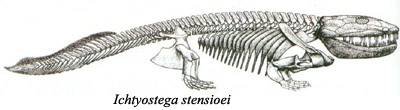
Udovi ihtiostege su prvi koji su direktno prilagođeni za hodanje po suvom tlu. Još uvek su prisutne razne riblje osobine: zglob između lobanje i kičme, oblik kičmenih kostiju, rep tipičan za rezoperke, lateralna linija… međutim, vodozemačke osobine su toliko izražene da se ihtiostega obično klasifikuje kao rani vodozemac.
Od ove tačke, rani vodozemci poput ihtiostege su se raširili svuda po svetu, i ubrzo se razdvojili na stotine raznih vrsta (na donjoj slici, primer je tulerpeton). Neke od ovih vrsta su postale preci kasnijih reptila, dok su druge postale preci današnjih sisara. Neke vrste su ostale vodozemci. A neke su čak ponovo napustile kopno, i vratile se potpuno vodenom životu (kao što je recimo bio slučaj sa krasigirinusom na donjoj slici; radi reference, ova životinja je bila preko metar i po dugačka, a njeni zakržljali udovi su bili dugi samo nekoliko centimetara). Evolucija nikada ne stoji, i adaptacije proizvode nove vrste u svim mogućim pravcima, sa svake moguće tačke…
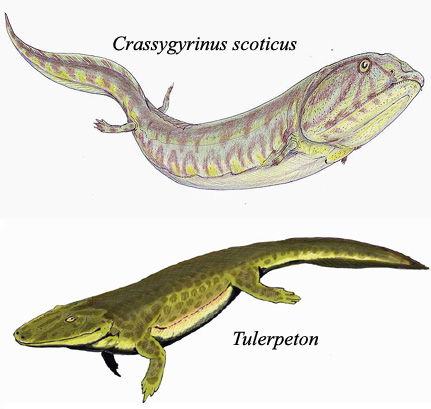
Autor rekonstrukcija: DiBgd
Ovaj gornji primer je sam po sebi dovoljna ilustracija fosilnih dokaza za evoluciju, kao i načina na koji se postepeno mogu razviti osobine koje izgledaju veoma kompleksno i daleko odvojeno od predačkih. Da bi smo ovu drugu poentu pokazali u potpunosti, pogledajmo još jedan primer – evoluciju modernih ptica.
Primer:Nastanak ptica. Danas većina ljudi smatra dinosauruse prosto jednom davno izumrlom vrstom reptila. U stvari, dinosaurusi su bili makar isto onoliko raznovrsni kao što su to sisari danas. Od gigantskih biljojeda, preko morskih i letećih životinja, pa sve do malih organizama veličine današnjih kokoški ili zečeva, reč dinosaurus obuhvata ogroman spektar vrsta.
Evolucija ptičjih pluća
Pored nastanka perja i krila, koji su objašnjeni u ovom tekstu, jedna od centralnih promena u nastanku ptica je nastanak kompleksnih ptičjih pluća. Pošto je ovo veoma komplikovan predmet, njime se nećemo baviti u ovom uvodnom tekstu, ali čitaoci koje on zanima mogu da pogledaju dodatni tekst na tu temu.
Jedna osnovna grupa dinosaurusa su takozvani teropodi, među koje spada i verovatno najpoznatiji dinosaurus, ogromni mesožder Tyrannosaurus rex. Svi teropodi su od svog zajedničkog pretka nasledili jednu grupu specifičnih osobina: hodali su na dve noge, imali su troprsta stopala, ključne kosti spojene u jednu (stručno zvanu furkula, a u narodu zvanu jadac), i kosti su im bile šuplje (radi brzine trčanja).

Fosilni skeleti dva teropoda, Herrasaurusa i Eoraptora.
Fotografija iz Severnoameričkog Muzeja Prastarog Života, autor Zak Tajrel (Zach Tirrell).
![]() Postoji više razloga zbog kojih je verovatno da su teropodi takođe bili makar delimično toplokrvne životinje; svakako, njihove fosile nalazimo u regionima koji su u njihovo vreme bili veoma hladni, što onemogućava život velikih hladnokrvnih životinja. Ono što, međutim, svakako znamo je da se kod teropoda veoma brzo pojavila još jedna osobina: izrasline na koži veoma slične zadebljanim dlačicama (shematski crtež A, desno), koje su omogućavale efikasnije održanje telesne toplote.
Postoji više razloga zbog kojih je verovatno da su teropodi takođe bili makar delimično toplokrvne životinje; svakako, njihove fosile nalazimo u regionima koji su u njihovo vreme bili veoma hladni, što onemogućava život velikih hladnokrvnih životinja. Ono što, međutim, svakako znamo je da se kod teropoda veoma brzo pojavila još jedna osobina: izrasline na koži veoma slične zadebljanim dlačicama (shematski crtež A, desno), koje su omogućavale efikasnije održanje telesne toplote.
Iz fosila, ne možemo tačno da znamo koje je poreklo ovih dlačica, ali strukturalna i genetska poređenja ukazuju da se verovatno radilo o produženju i zadebljanju senzornih dlačica koje mnogi reptili imaju između krljušti (što poboljšava čulo dodira). Primer jednog fosila na kome ovo jasno možemo da vidimo je ostatak male životinje zvane sinosauropteriks:

Fosil Sinosauropteryx prima, u Muzeju Unutrašnje Mongolije. Autor Olai Ose.
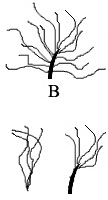 Analiza iz januara 2010-te je pokazala naizmenično prisustvo pigmenata i pigmentnih ćelija u okviru fosila sinosauropteriksa, što znači da su njegove dlake bile raznobojne, i da je rep skoro sigurno bio prugast. Ove “dlačice” su najraniji oblik izraslina kakve vidimo i na mnogim drugim fosilima. Recimo, rani srodnici i preci tiranosaurida su imali slično perje kao i sinosauropteriks.
Analiza iz januara 2010-te je pokazala naizmenično prisustvo pigmenata i pigmentnih ćelija u okviru fosila sinosauropteriksa, što znači da su njegove dlake bile raznobojne, i da je rep skoro sigurno bio prugast. Ove “dlačice” su najraniji oblik izraslina kakve vidimo i na mnogim drugim fosilima. Recimo, rani srodnici i preci tiranosaurida su imali slično perje kao i sinosauropteriks.
No, stvari nisu dugo ostale iste. Kod jedne kasnije grupe teropoda, takozvanih terizinosaurusa (ogromnih životinja sa velikim kandžama na prednjim udovima), možemo da vidimo kako se početne dlačice razvijaju u nekoliko sličnih struktura. U različitim vrstama (ili na različitim delovima tela iste vrste) vidimo višestruke dlačice, debele dlake koje se šire u više dlačica na vrhu, i debele centralne dlačice sa manjim dlačicama sa strane (shematski crtež B, desno). Primer ovakve životinje je notronikus, biljožderni dinosaur visok šest metara i težak oko jedne tone:

Nothronychus mckinleyi, rekonstrukcija izgleda.
Autor: Arthur Weasley.
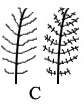 Naravno, i ovo je samo primer jednog koraka. Niz od nekoliko desetina dinosaurskih fosila pokazuje dalji razvoj perja. U varijaciji na gornju shemu “B”, centar pera se stabilizuje, dlačice sa strane postaju paralelnije, i u trećem koraku nastaje ukrštanje koje proizvodi prepoznatljivo pero (shema C, desno).
Naravno, i ovo je samo primer jednog koraka. Niz od nekoliko desetina dinosaurskih fosila pokazuje dalji razvoj perja. U varijaciji na gornju shemu “B”, centar pera se stabilizuje, dlačice sa strane postaju paralelnije, i u trećem koraku nastaje ukrštanje koje proizvodi prepoznatljivo pero (shema C, desno).
Važno je ovde razumeti da mi ne govorimo samo o precima današnjih ptica. Mnoge ove vrste su bili prosto pernati dinosaurusi, čiji potomci su izumrli nakon Jukatanskog meteorskog udara, i kojih više danas nigde nema. Recimo, jedan lep primer pernatog dinosaurusa koji nije ptica niti je predak ptica je kaudipteriks:
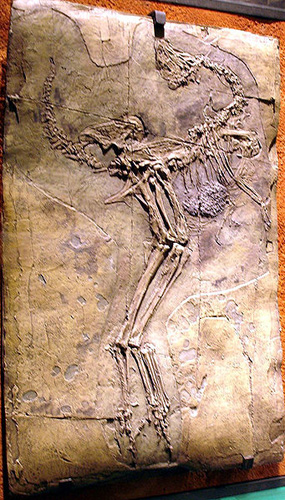
Caudipteryx fosil, očuvan toliko dobro da imamo i fosil sadržaja stomaka u trenutku smrti.
Fild muzej, Čikago. Fotografija: Yo Hibino
![]() Kaudipteriks je imao telo prekriveno “razgranatim perolikim dlačicama”, sličnim onim na shemi C. Ali istovremeno, imao je kratko prepoznatljivo perje (shematski crtež D) na repu, prednjim udovima i na nogama. Možemo da nagađamo da je ovo perje verovatno korišćeno u svrhe privlačenja ženki, ili se širilo da bi životinja izgledala veća pred napadačima; no, pošto se ponašanje životinja ne fosilizuje, ne možemo ovo znati sigurno. Ono što možemo znati je da ovo nisu krila, i da nisu korišćena za let – kaudipteriks nije ptica, nema letne mišiće, a perje nije ni dovoljno dugačko niti prilagođeno za let.
Kaudipteriks je imao telo prekriveno “razgranatim perolikim dlačicama”, sličnim onim na shemi C. Ali istovremeno, imao je kratko prepoznatljivo perje (shematski crtež D) na repu, prednjim udovima i na nogama. Možemo da nagađamo da je ovo perje verovatno korišćeno u svrhe privlačenja ženki, ili se širilo da bi životinja izgledala veća pred napadačima; no, pošto se ponašanje životinja ne fosilizuje, ne možemo ovo znati sigurno. Ono što možemo znati je da ovo nisu krila, i da nisu korišćena za let – kaudipteriks nije ptica, nema letne mišiće, a perje nije ni dovoljno dugačko niti prilagođeno za let.
Dakle, perje nije nastalo radi leta, niti je evoluiralo kod letećih organizama. Umesto toga, fosili nam pokazuju da je ono nastalo kod dinosaurusa, najverovatnije iz istih razloga iz koga i sisarska dlaka: radi očuvanja telesne toplote. Mi ne možemo biti sigurni da li su dinosaurusi bili toplokrvni ili hladnokrvni (mada možemo naći neke argumente za toplokrvnost, oni nisu sigurni); međutim, u oba slučaja, perje omogućava bolje održanje toplote i dužu aktivnost u hladnim uslovima, što je snažan selektivni pritisak.
![]() Razvoj leta se desio sekundarno: kada su već postojali pernati dinosaurusi, neki od njih su počeli da lete. Kako se tačno razvio sam let, u smislu ponašanja životinje, je jako teško zaključiti na osnovu fosila.
Razvoj leta se desio sekundarno: kada su već postojali pernati dinosaurusi, neki od njih su počeli da lete. Kako se tačno razvio sam let, u smislu ponašanja životinje, je jako teško zaključiti na osnovu fosila.
No, možemo da nađemo neke tačke za orjentaciju. Recimo, zbog aerodinamike, pero letećih životinja mora biti asimetrično (shematski crtež E). Pošto je malo verovatno da ovakva pera mogu nastati bez specifičnog selektivnog pritiska, fosili koji poseduju asimetrično perje su verovatno ostaci životinja koje su makar bile u stanju da jedre. Uz to, možemo pratiti nastanak osobina koje su usko povezane sa letom (snažnije kosti za vezivanje letnih mišića, veće i snažnije perje, i mnoge druge osobine).
Pernati teropodi sa perjem vrste “D” su imali to perje na repu i na sva četiri uda. Prvi organizmi koji su razvili asimetrično perje “E” su imali isti raspored. Dobar primer ovoga je dinosauroptica zvana mikroraptor:
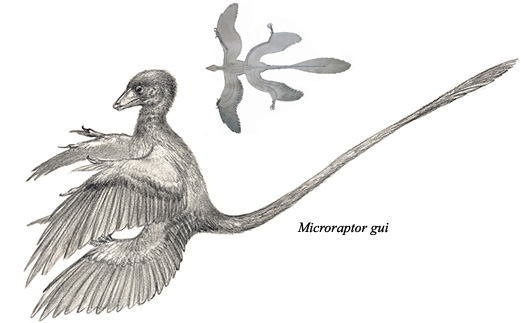
Xu, X., Zhou, Z., Wang, X. et al. Four-winged dinosaurs from China. Nature 421, 335–340 (2003).
Ovi prvi leteći dinosaurusi (ne može se govoriti još o pticama) su bili veličine omanjeg goluba, i mala veličina im je olakšavala korišćenje perja za jedrenje i letenje. Oni su takođe nasledili početne osobine terapoda kroz celu stazu nasleđa: svi terapodi, od tiranosaurusa i velociraptora, preko ovih prvi letača, pa sve do današnjih ptica i dalje imaju ključne kosti povezane u jadac, troprste noge, i lake, šuplje kosti. Mnoge osobine današnjih ptica i dalje nedostaju, što se vidi na primeru čuvenog arheopteriksa, koji se smatra tačkom na kojoj pticoliki dinosaurusi prelaze u dinosaurolike ptice:

Archaeopteryx lithographica, Berlinski primerak.
Fotografija: H. Raab.
Mada se arheopteriksova sposobnost leta vidi na osnovu mnogobrojnih adaptacija i organizacije perja u prava krila, on je i dalje velikim delom dinosaur. Kičma je još uvek ista kao kod prethodnih pernatih dinosaurusa, i dalje ima vilicu sa zubima, dugačak i koščat rep, ravnu grudnu kost, i kandže na krilima.
Ove osobine se polako, korak po korak menjaju u daljim fosilima. Zubi polako nestaju (fosil rane ptice zvane Confuciusornis je prvi kod koga se vilica izdužuje u pravi kljun), rep se skraćuje, grudi postaju tipično ptičje a grudni mišići sve snažniji. Iz ove osnovne grupe prvih letećih, pernatih dinosaurusa, razvijaju se dve grupe životinja. Jedna od njih, Enantiornithes, se proširila na mnogo vrsta, koje su sve nestale zajedno sa dinosaurusima; druga grupa, Euornithes, je preživela ovaj pomor, i njihovi potomci su naše današnje ptice.
Dinosaursko poreklo ptica je, naravno, podržano i podacima koji su potpuno nezavisni od fosilnih. Genom ptica je u ugnježdenoj hijerarhiji najbliži genomu reptila. Ptice i dalje sadrže u svom genomu ostatke, pseudogene, iz reptilskog doba; grupa naučnika je, recimo, kod kokošaka reaktivirala jednu tu ugašenu grupu (ništa nisu dodavali, samo su aktivirali postojeće ugašene gene), i dobili su piliće sa vilicama i zubima umesto kljunova.
Uspešno je nađena i jedna potpuno direktna genetska veza. Iz tri veoma dobro očuvana fosila dinosaurusa, tri grupe naučnika su uspele da rekonstruišu mineralizovane ostatke vezivnih proteina (koji su među najotpornijim supstancama u živom svetu), i da sekvence tih proteina uporede sa modernim živim bićima. Po tom poređenju, opet, grupa koja je danas najbliža dinosaurusima su upravo ptice.
Za kraj ovog teksta, pogledajmo jedan završni primer, koji lepo pokazuje kako evolucija adaptira postojeće osobine, i kako takve adaptacije mogu proizvesti veoma kompleksne rezultate.
Primer:Slušni aparat sisara. Ako počnemo da istražujemo anatomiju današnjih životinja, naćićemo mnoge veoma kompleksne strukture. Standardan primer za ovo je oko, čija evolucija je objašnjena na ovom sajtu u celom posebnom tekstu. Pošto ovde govorimo o prelaznim fosilima, možemo baciti pogled na jedan takođe veoma komplikovan sistem, sisarski slušni sistem:
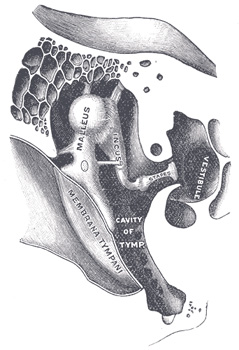
U okviru ovog sistema se nalaze tri sićušne kosti, zvane “čekić,” “nakovanj” i “uzengija.” Jedan minijaturni mišić povezuje čekić i nakovanj, omogućavajući kontrolu prenosa zvuka. Zvuk se prenosi sa bubne opne (na dnu ušnog kanala), kroz ove tri kosti, u specijalan aparat u okviru koga vibracije zvuka pokreću specijalne “dlakave ćelije”.
Ovaj sistem je odličan primer velike kompleksnosti koja može nastati u okviru živog carstva. Dlakave ćelije i osnovni unutrašnji sistem sluha je prisutan kod riba, i preneo se kroz niz nasleđa koji je opisan u prvom primeru iznad. Ali kako ceo ovaj ostali sistem može nastati kroz evolutivne procese?
Na sreću, deo lobanje koji okružuje slušni aparat je među najtvrđim i najotpornijim kostima u telu (samo su zubi čvršći), a kosti iz kojih je on evoluirao su kosti vilice (takođe među najčvršćim i najotpornijim). Ovo znači da je preživelo mnogo prelaznih fosila, na osnovu kojih nam je ova evolutivna staza poznata veoma detaljno.
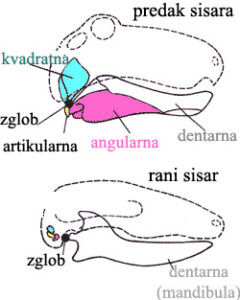 Da bi smo razumeli nastanak slušnog sistema, moramo da vidimo od čega on počinje. Najraniji preci današnjih sisara su potomci onih amfibijskih organizama koje smo opisali u prvom primeru. Ovi organizmi su imali kompleksnu donju vilicu, koja se u prednjem delu sastojala od dentarne kosti (u koju su bili usađeni zubi) i angularne kosti sa strane vilice. Zglob vilice se nalazio između takozvane kvadratne i artikularne kosti; zvučne vibracije su se prenosile uz vilicu do kvadratne kosti, koja ih je prenosila u slušni sistem (veoma primitivan i zasnovan na prenosu vibracija kroz vestibulu).
Da bi smo razumeli nastanak slušnog sistema, moramo da vidimo od čega on počinje. Najraniji preci današnjih sisara su potomci onih amfibijskih organizama koje smo opisali u prvom primeru. Ovi organizmi su imali kompleksnu donju vilicu, koja se u prednjem delu sastojala od dentarne kosti (u koju su bili usađeni zubi) i angularne kosti sa strane vilice. Zglob vilice se nalazio između takozvane kvadratne i artikularne kosti; zvučne vibracije su se prenosile uz vilicu do kvadratne kosti, koja ih je prenosila u slušni sistem (veoma primitivan i zasnovan na prenosu vibracija kroz vestibulu).
Ovi organizmi su imali uzengiju, ali je ona bila relativno velika i stajala je sa strane moždane šupljine. Takođe, ušna duplja je bila otvorena, i nije bilo bubne opne. Ukupno, zbog nasleđenog ribljeg slušnog sistema, ove životinje su bile skoro gluve – čule su samo zvuke najnižih frekvencija.
Možemo ovde da uočimo i jedan dodatni problem: vilični zglob današnjih sisara se nalazi između prednje kosti vilice (mandibule, ekvivalentne dentarnoj) i kostiju lobanje. Na prvi pogled ovo zahteva da potpuno oformljen zglob nestane sa jednog mesta, i da se odjednom pojavi na potpuno drugom! Prelazni fosili, kao što ćemo videti, daju odgovor i na ovo pitanje….
Slab sluh prvih amfibijskih organizama nije predstavljao preterano veliku prepreku, pošto su sve druge kopnene životinje tog doba patile od istog problema. No, postojao je snažan selektivni pritisak za razvoj boljeg sluha, pošto svako poboljšanje nosi sa sobom i povećanu šansu preživljavanja. U precima sisara, mutacije koje povećavaju dentatnu kost, a smanjuju ostale pomenute kosti, automatski proizvode bolji prenos viših frekvencija. Svaka pojedinačna mutacija, svaka mala promena, nosi sa sobom malo poboljšanje sluha.
Otud, nije iznenađujuće što u fosilnom zapisu vidimo postepeno smanjenje kvadratne, artikularne i angularne kosti, uz istovremeno povećanje dentatne, koja se širi u stranu i unazad. Ove promene se odigravaju kroz seriju veoma malih koraka, u veoma postepenoj evoluciji. U ovom povlačenju, smanjenje kvadratne kosti menja položaj uzengije, koja postaje vezana za unutrašnji slušni aparat.
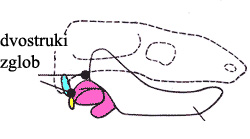 U toku ovog smanjenja kostiju, dolazimo do trenutka kada rastuća dentarna kost (koju sada već možemo da zovemo sisarskim imenom: mandibula) dođe u kontakt sa kostima lobanje. Brojni fosili predaka sisara poznatih kao morganukodoni pokazuje šta se desilo: na novoj tački kontakta između dve kosti, iz neophodnosti se pojavio novi zglob. Time imamo životinju sa dvostrukim viličnim zglobom, i rešenje gorepomenutog problema: od ove tačke nadalje, prednji “sisarski” zglob postaje sve razvijeniji, dok zadnji “reptilski” zglob polako nestaje.
U toku ovog smanjenja kostiju, dolazimo do trenutka kada rastuća dentarna kost (koju sada već možemo da zovemo sisarskim imenom: mandibula) dođe u kontakt sa kostima lobanje. Brojni fosili predaka sisara poznatih kao morganukodoni pokazuje šta se desilo: na novoj tački kontakta između dve kosti, iz neophodnosti se pojavio novi zglob. Time imamo životinju sa dvostrukim viličnim zglobom, i rešenje gorepomenutog problema: od ove tačke nadalje, prednji “sisarski” zglob postaje sve razvijeniji, dok zadnji “reptilski” zglob polako nestaje.
Kroz dalju minijaturizaciju kostiju, sluh postaje oštar i u visokim frekvencijama, i nastaje moderni slušni sistem.
Sad, kada smo sve ovo opisali rečima, pogledajmo i dijagram. Na sledećoj slici su sa fosila precrtane sheme vilica sisarskih predaka, poređane od najstarijih (na dnu) do najmlađih (na vrhu). Artikularna kost (na slici označena sa Ar) se polako smanjuje i postaje čekić (malleus, Ma); kvadratna kost (Q) se takođe smanjuje i postaje nakovanj (incus, I). Angularna kost (An) je postala spoljni deo aparata, koji se kod današnjih sisara zove annulus tympani (označena sa ty).
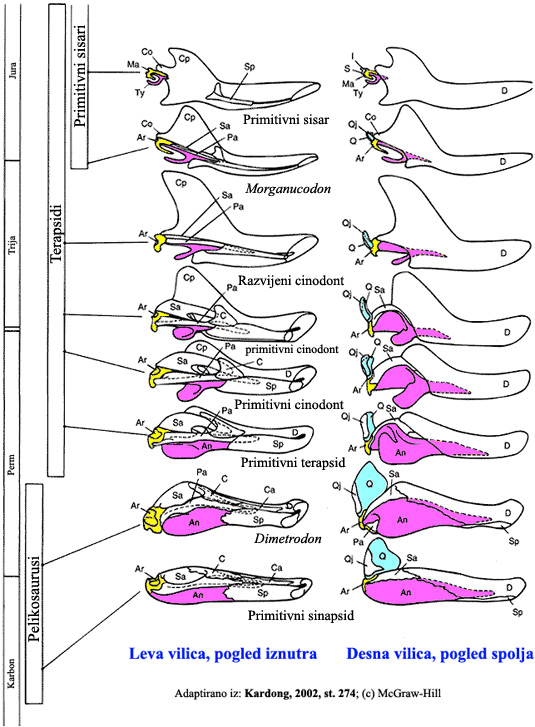
Evolucija naizgled nemoguće kompleksnog sistema je na ovaj način ilustrovana u koracima koji su sasvim u rangu veličine promena kakve danas posmatramo u prirodi.
Za one čitaoce koji imaju dodatna pitanja o procesima fosizilacije i načinima na koje nastaju ovakvi fosili, ispod je dat jedan dodatan kratak pregled. Za one koji žele da vide još primera prelaznih fosila, tu je naravno ceo tekst na tu temu. A za sve ostale, pre nego što zaključimo ovaj tekst, u sledećem poglavlju ćemo pogledati još nekoliko sporednih pokazatelja evolucije i zajedničkog nasleđa.
Proces fosilizacije i razlozi za retkost fosila
Ako se tkivo životinje nađe u odgovarajućim uslovima, u kojima nema daljeg razlaganja kroz dejstvo mikroorganizama (ili u kojima je to razlaganje veoma usporeno), može doći do njegove mineralizacije. Ovo je proces u toku koga se minerali iz okolnog zemljišta i vode polako nakupljaju na površini i unutar tkiva, proizvodeći kristalnu strukturu drugačiju od okolnih sedimenata. Nakon dugog perioda vremena, organski materijali će biti potpuno razoreni, i zamenjeni mineralima: ali ti minerali će imati oblik mineralizovanog tkiva. Na ovaj način, u sedimentnim naslagama povremeno možemo naći fosile, mineralizovane ostatke biljaka, životinja, pa čak povremeno i mikroorganizama.
Uzimajući u obzir dugu istoriju Zemlje, i broj živih bića koji je na njoj postojao u toku te istorije, fosili takvih bića bi trebali da budu veoma česta stvar. Otud, možemo se pitati zašto su fosilni ostaci retki, i zašto nemamo fosile svih životinja koje su ikada postojale. Odgovor na ovo pitanje je relativno jednostavan: ostaci živih bića su veoma česti (ima ih na trilione tona), i među njima verovatno ima bar ponekih ostataka većine vrsta koje su postojale. Problem je što ti ostaci nisu u jednom komadu…
Ostaci biljaka i životinja su odličan izvor hrane drugim organizmima i mikoroorganizmima, zbog čega nakon smrti skoro svi takvi ostaci bivaju prosto pojedeni (kada mikroorganizmi “vare” ostatke drugih organizama, mi to zovemo “truljenje”). U takvim slučajevima, ono što dugoročno ostaje su samo teže svarljivi delovi tela.
Tako da danas imamo mnogobrojne naslage koje se sastoje od hiljada kubnih kilometara nagomilanih i mineralizovanih komadića kostiju pomešanih sa vulkanskim peskom, ostataka naslaganih školjki, pa čak i kilometarske formacije koje se dobrim delom sastoje od ribljih zuba (zubi i vilice su među najtvrđim kostima, i otud su njihovi fosili praktično bezbrojni).
Geološki stub
Za dalje informacije o geološkim slojevima i o količinama nagomilane organske materije, pogledajte tekst sa primerom jednog geološkog stuba.
Veoma je retko da čitave kosti ostanu u komadu. Još ređe je da se to desi sa celim delom tela (recimo, celim udom, ili celom lobanjom). A za očuvanje celog tela, potrebni su fantastično retki uslovi – toliko retki da većina vrsta na Zemlji nikada neće ostaviti ni jedan jedini kompletan fosil.
Postoje, naravno, izuzeci. Pojedinačno najveći su diatomi, jednoćelijske morske životinje koje svoje ćelije okružuju mineralnim zidom. Nakon smrti, organski deo diatoma biva uništen (ili pojeden), ali ove “ćelijske školjke” ostaju manje-više cele, i polako se talože na dnu. Ono što mi zovemo kreda nije ništa drugo do upravo takog ovakvih jednoćelijskih kostura; veliki deo krečnjaka takođe spada u ovu klasu, i postoje čitave planine koje se sastoje od naslaga ostataka ovih organizama.
Bića sa školjkama, koje su sastavljene od minerala kalcijum-karbonata (i otud ne mogu da se pojedu) ostavljaju gomile fosila svojih školjki ili oklopa. Močvare ili veoma mirna, duboka jezera, takođe predstavljaju odlična mesta za fosilizaciju, pošto ostaci životinje koji padnu na dno često mogu biti zatrpani u anoksičnom mulju (blatu sa malo kiseonika), gde su procesi truljenja spori, a kosti se mineralizuju veoma brzo. Za očuvanje drugih karakteristika organizma – mekog tkiva, ili perja, dlake i sličnih stvari – potrebni su još ređi i specifičniji uslovi.
Povrh svega ovoga, sedimenti su uvek izloženi tektonskim procesima, i procesima erozije. Na mestima subdukcije tektonskih ploča, sedimentne stene bivaju odnesene u magmu zajedno sa vulkanskim. Takođe, ako sloj u kome se fosil nalazi postane izložen površini, on će biti spran i uništen u roku od nekoliko decenija.
Zbog svega ovoga, mi nikada nećemo imati potpunu fosilnu istoriju sveta. Suviše vrsta se nikada nije fosilizovalo, suviše fosila je već uništeno erozijom, suviše ih se nalazi razbacano u stenju na dubinama koje se mere kilometrima, i suviše nikada neće biti nađeno. Ali mi i pored toga imamo na desetine hiljada prelaznih fosila, što imamo je sasvim dovoljno da pokaže kako se život razvijao kroz vreme, i kako je nastala raznovrsnost koju vidimo danas.
Lobanje ljudskih predaka
Fosilni dokazi o evoluciji čoveka su detaljno objašnjeni u tekstu na tu temu; ali za one koji su imali strpljenja da pročitaju celo ovo poglavlje, kao malu dodatnu nagradu nudimo jednu uporednu sliku. Pogledajte sami, i vidite da li možete da odlučite gde prestaju hominini, a gde počinje čovek…
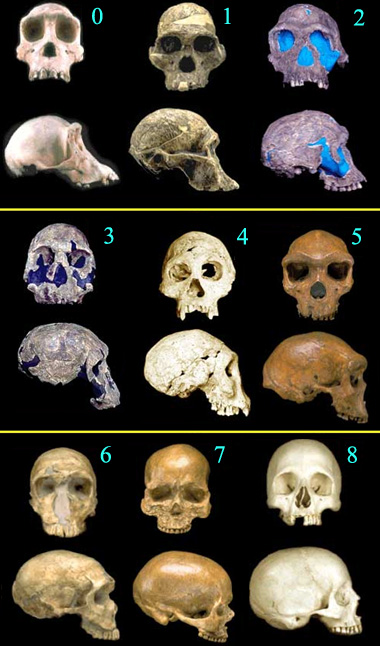
- Australopithecus africanus, fosil STS 5, starost 2,6 miliona godina
- Homo habilis, fosil KNM-ER 1813, starost 1,9 miliona godina
- Homo rudolfensis, fosil KNM-ER 1470, starost 1,8 miliona godina
- Homo erectus, fosil D2700, starost 1,75 miliona godina
- Homo heidelbergensis, starost 300-125 hiljada godina
- Homo sapiens neanderthalensis, fosil La Ferrassie 1, starost 70 hiljada godina
- Homo sapiens sapiens, fosil Kro-Manjon I, starost 30 hiljada godina
- Moderni čovek,Homo sapiens sapiens, danas
Radi poređenja, pod brojem 0 je dodata lobanja modernog šimpanze (Pan troglodytes). Kliknite ovde za veću i kompletniju kolekciju lobanja prelaznih fosila ljudskih predaka. Sve fotografije su © 2000 Smithsonian Institution.
[prethodno poglavlje][nazad na vrh strane][sledeće poglavlje]